

Relationships between Slc1a5 and Osteoclastogenesis
- PMID: 34301346
- PMCID: PMC8384000
- DOI: 10.30802/AALAS-CM-21-000012
Relationships between Slc1a5 and Osteoclastogenesis
Abstract
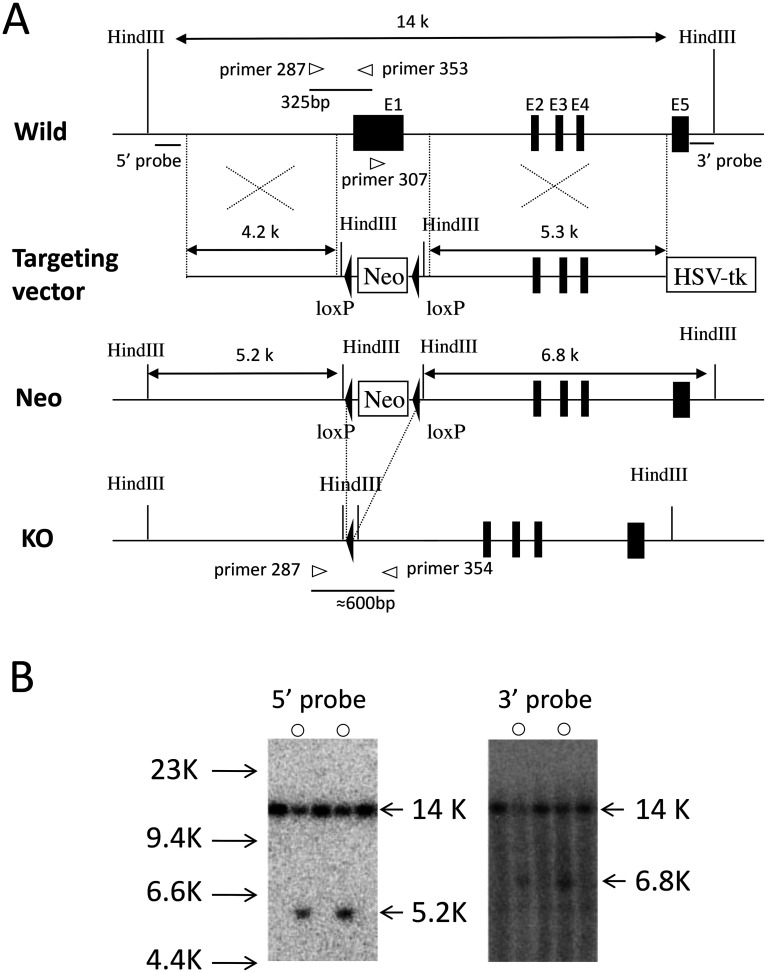
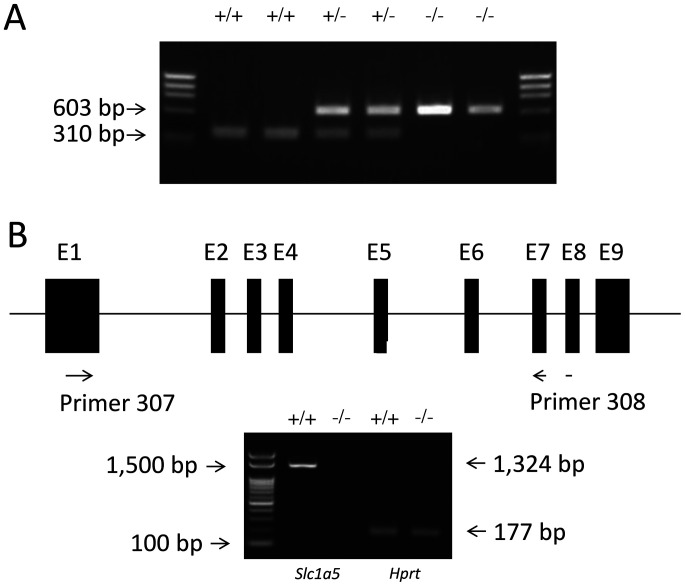
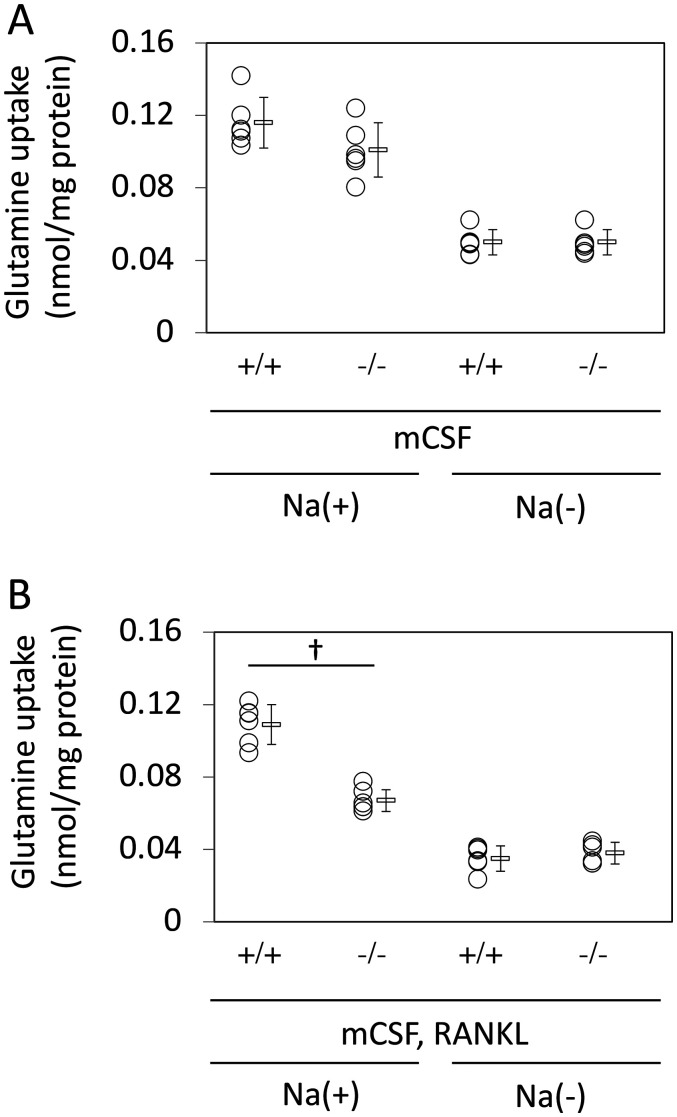
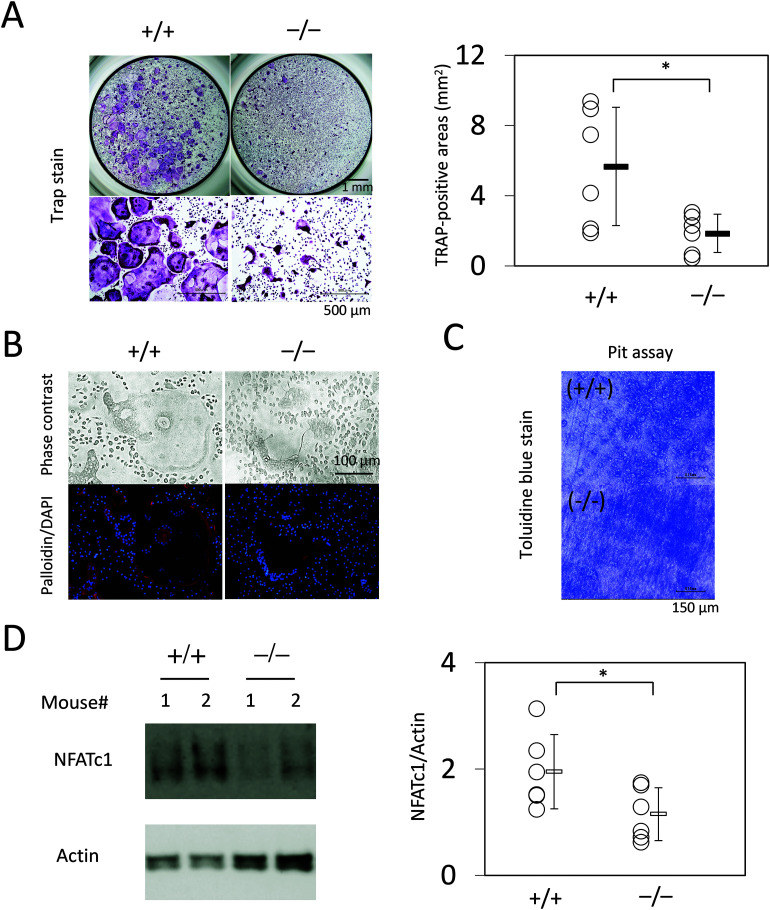
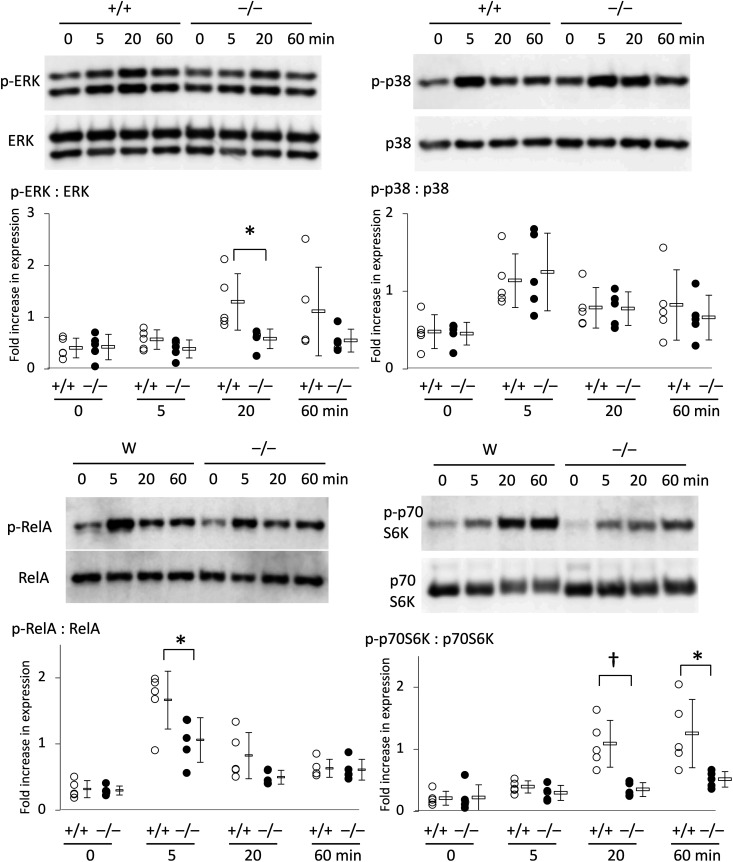
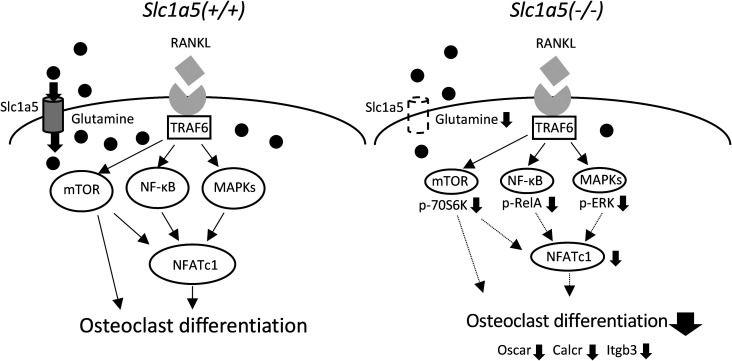
Slc1a5 (ASCT2) encodes a small neutral amino-acid exchanger and is the most well-studied glutamine transporter in cancer cells. To investigate the role of Slc1a5 in osteoclastogenesis, we developed Slc1a5-deficient mice by using a conventional gene-targeting approach. The Slc1a5-/- mice showed no obvious abnormalities in growth. Glutamine uptake was assessed in Slc1a5+/+ and Slc1a5-/- bone marrow cells stimulated with RANKL. The rate of glutamine uptake in Slc1a5-/- bone marrow cells was reduced to 70% of that of cells from Slc1a5+/+ bone marrow. To confirm the involvement of Slc1a5 in osteoclast formation, bone marrow cells derived from Slc1a5+/+ or Slc1a5-/- mice were stimulated with RANKL and macrophage colony-stimulating factor and stained with tartrate-resistant acid phosphatase. The bone resorption activity and actin ring formation of stimulated cells were measured. The formation of multinucleated osteoclasts in bone marrow cells isolated from Slc1a5-/- mice was severely impaired compared with those from Slc1a5+/+ mice. RANKL-induced expression of ERK, NFκB, p70S6K, and NFATc1 was suppressed in Slc1a5-/- osteoclasts. These results show that Slc1a5 plays an important role in osteoclast formation.
Figures

Similar articles
-
Kalkitoxin Reduces Osteoclast Formation and Resorption and Protects against Inflammatory Bone Loss.Int J Mol Sci. 2021 Feb 25;22(5):2303. doi: 10.3390/ijms22052303. Int J Mol Sci. 2021. PMID: 33669069 Free PMC article.
-
Metabolic regulation of osteoclast differentiation and function.J Bone Miner Res. 2013 Nov;28(11):2392-9. doi: 10.1002/jbmr.1976. J Bone Miner Res. 2013. PMID: 23661628
-
Deletion of CD74, a putative MIF receptor, in mice enhances osteoclastogenesis and decreases bone mass.J Bone Miner Res. 2013 Apr;28(4):948-59. doi: 10.1002/jbmr.1787. J Bone Miner Res. 2013. PMID: 23044992 Free PMC article.
-
Constant hypoxia inhibits osteoclast differentiation and bone resorption by regulating phosphorylation of JNK and IκBα.Inflamm Res. 2019 Feb;68(2):157-166. doi: 10.1007/s00011-018-1209-9. Epub 2019 Jan 2. Inflamm Res. 2019. PMID: 30604211
-
Osteoclast differentiation by RANKL and OPG signaling pathways.J Bone Miner Metab. 2021 Jan;39(1):19-26. doi: 10.1007/s00774-020-01162-6. Epub 2020 Oct 20. J Bone Miner Metab. 2021. PMID: 33079279 Review.
Cited by
-
Mechanistic insights into bone destruction in multiple myeloma: Cellular and molecular perspectives.J Bone Oncol. 2025 Mar 4;51:100668. doi: 10.1016/j.jbo.2025.100668. eCollection 2025 Apr. J Bone Oncol. 2025. PMID: 40124903 Free PMC article. Review.
-
Novel Insights into Osteoclast Energy Metabolism.Curr Osteoporos Rep. 2023 Dec;21(6):660-669. doi: 10.1007/s11914-023-00825-3. Epub 2023 Oct 10. Curr Osteoporos Rep. 2023. PMID: 37816910 Free PMC article. Review.
-
Role of amino acid metabolism in osteoporosis: Effects on the bone microenvironment and treatment strategies (Review).Mol Med Rep. 2025 Aug;32(2):212. doi: 10.3892/mmr.2025.13577. Epub 2025 May 26. Mol Med Rep. 2025. PMID: 40417875 Free PMC article. Review.
-
Amino acid metabolism in skeletal cells.Bone Rep. 2022 Sep 8;17:101620. doi: 10.1016/j.bonr.2022.101620. eCollection 2022 Dec. Bone Rep. 2022. PMID: 36120644 Free PMC article.
References
-
- Bothwell PJ, Kron CD, Wittke EF, Czerniak BN, Bode BP.2018.Targeted suppression and knockout of ASCT2 or LAT1 in epithelial and mesenchymal human liver cancer cells fail to inhibit growth.Int J Mol Sci 19:1–25.10.3390/ijms19072093. - DOI - PMC - PubMed
-
- Bröer A, Gauthier-Coles G, Rahimi F, van Geldermalsen M, Dorsch D, Wegener A, Holst J, Bröer S.2019.Ablation of the ASCT2 (SLC1A5) gene encoding a neutral amino acid transporter reveals transporter plasticity and redundancy in cancer cells.J Biol Chem 294:4012–4026.10.1074/jbc.RA118.006378. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
