

Reinforcement learning links spontaneous cortical dopamine impulses to reward
- PMID: 34302743
- PMCID: PMC8605927
- DOI: 10.1016/j.cub.2021.06.069
Reinforcement learning links spontaneous cortical dopamine impulses to reward
Abstract
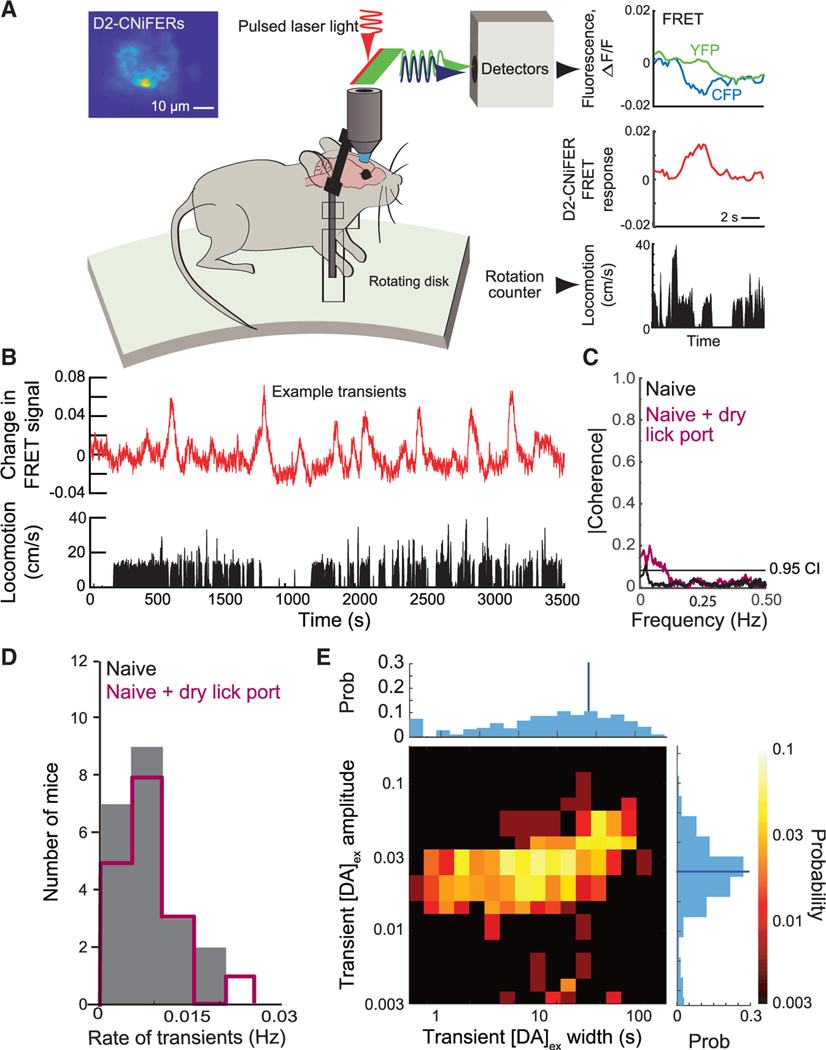
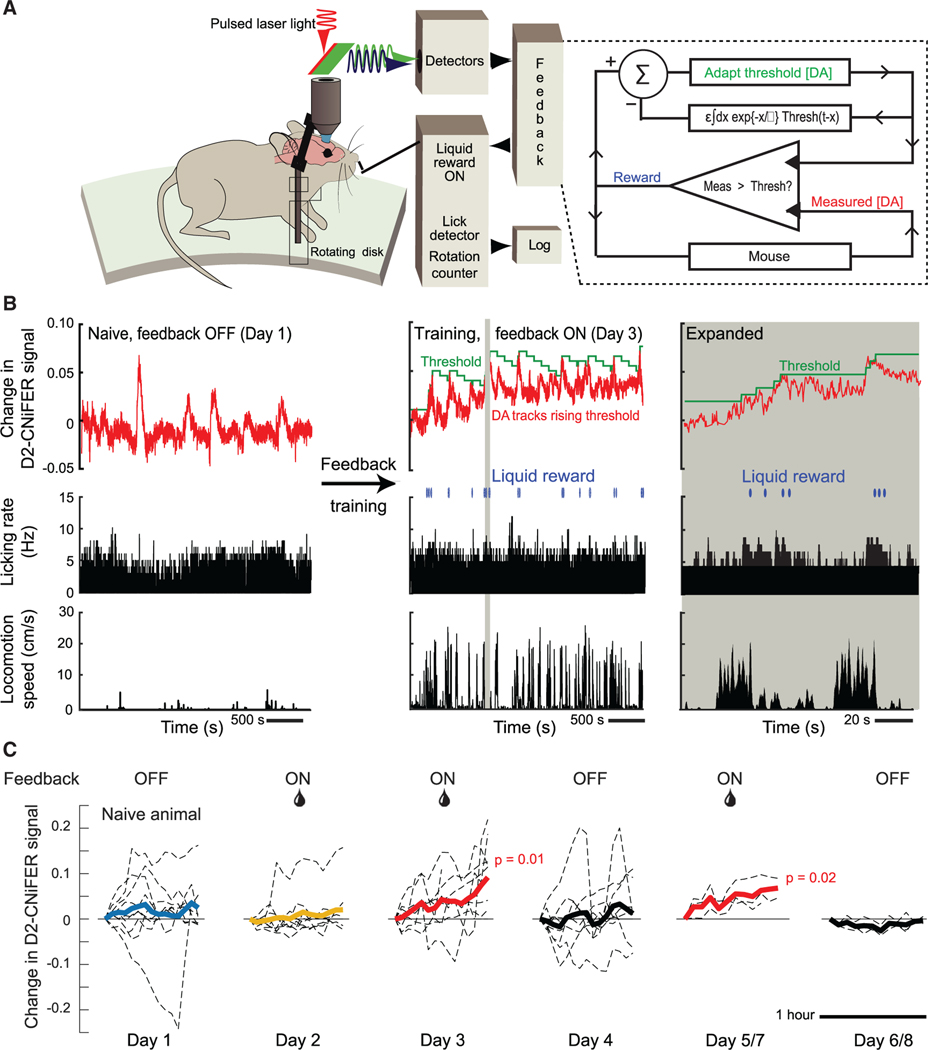
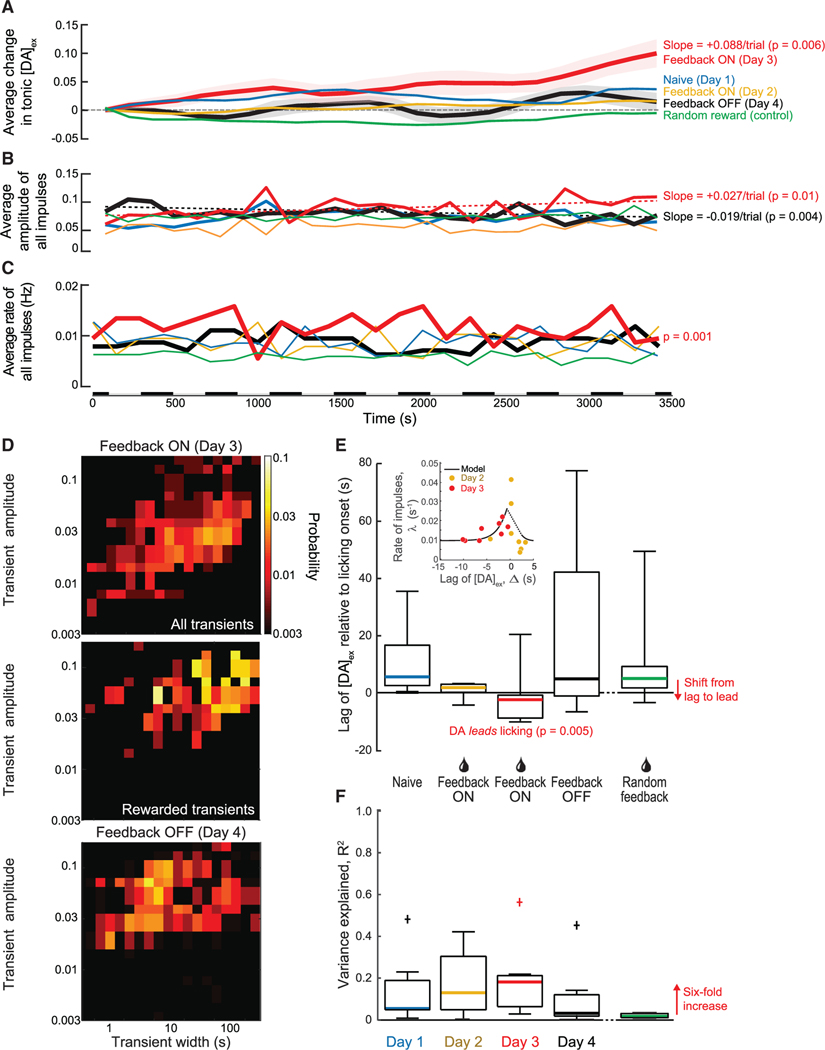
In their pioneering study on dopamine release, Romo and Schultz speculated "...that the amount of dopamine released by unmodulated spontaneous impulse activity exerts a tonic, permissive influence on neuronal processes more actively engaged in preparation of self-initiated movements...."1 Motivated by the suggestion of "spontaneous impulses," as well as by the "ramp up" of dopaminergic neuronal activity that occurs when rodents navigate to a reward,2-5 we asked two questions. First, are there spontaneous impulses of dopamine that are released in cortex? Using cell-based optical sensors of extrasynaptic dopamine, [DA]ex,6 we found that spontaneous dopamine impulses in cortex of naive mice occur at a rate of ∼0.01 per second. Next, can mice be trained to change the amplitude and/or timing of dopamine events triggered by internal brain dynamics, much as they can change the amplitude and timing of dopamine impulses based on an external cue?7-9 Using a reinforcement learning paradigm based solely on rewards that were gated by feedback from real-time measurements of [DA]ex, we found that mice can volitionally modulate their spontaneous [DA]ex. In particular, by only the second session of daily, hour-long training, mice increased the rate of impulses of [DA]ex, increased the amplitude of the impulses, and increased their tonic level of [DA]ex for a reward. Critically, mice learned to reliably elicit [DA]ex impulses prior to receiving a reward. These effects reversed when the reward was removed. We posit that spontaneous dopamine impulses may serve as a salient cognitive event in behavioral planning.
Keywords: biophysical modeling; brain machine interface; classical conditioning; feedback; foraging; neuromodulation; stochastic dynamics; two-photon microscopy.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Tonic or Phasic Stimulation of Dopaminergic Projections to Prefrontal Cortex Causes Mice to Maintain or Deviate from Previously Learned Behavioral Strategies.J Neurosci. 2017 Aug 30;37(35):8315-8329. doi: 10.1523/JNEUROSCI.1221-17.2017. Epub 2017 Jul 24. J Neurosci. 2017. PMID: 28739583 Free PMC article.
-
Dopamine errors drive excitatory and inhibitory components of backward conditioning in an outcome-specific manner.Curr Biol. 2022 Jul 25;32(14):3210-3218.e3. doi: 10.1016/j.cub.2022.06.035. Epub 2022 Jun 24. Curr Biol. 2022. PMID: 35752165
-
Responses of monkey dopamine neurons during learning of behavioral reactions.J Neurophysiol. 1992 Jan;67(1):145-63. doi: 10.1152/jn.1992.67.1.145. J Neurophysiol. 1992. PMID: 1552316
-
Predictive reward signal of dopamine neurons.J Neurophysiol. 1998 Jul;80(1):1-27. doi: 10.1152/jn.1998.80.1.1. J Neurophysiol. 1998. PMID: 9658025 Review.
-
Neural control of dopamine neurotransmission: implications for reinforcement learning.Eur J Neurosci. 2012 Apr;35(7):1115-23. doi: 10.1111/j.1460-9568.2012.08055.x. Eur J Neurosci. 2012. PMID: 22487041 Review.
Cited by
-
Closed-loop experiments and brain machine interfaces with multiphoton microscopy.Neurophotonics. 2024 Jul;11(3):033405. doi: 10.1117/1.NPh.11.3.033405. Epub 2024 Feb 19. Neurophotonics. 2024. PMID: 38375331 Free PMC article. Review.
-
Genetically encoded sensors illuminate in vivo detection for neurotransmission: Development, application, and optimization strategies.IBRO Neurosci Rep. 2025 Mar 13;18:476-490. doi: 10.1016/j.ibneur.2025.03.003. eCollection 2025 Jun. IBRO Neurosci Rep. 2025. PMID: 40177704 Free PMC article.
-
Three Water Restriction Schedules Used in Rodent Behavioral Tasks Transiently Impair Growth and Differentially Evoke a Stress Hormone Response without Causing Dehydration.eNeuro. 2021 Dec 14;8(6):ENEURO.0424-21.2021. doi: 10.1523/ENEURO.0424-21.2021. Print 2021 Nov-Dec. eNeuro. 2021. PMID: 34815297 Free PMC article.
-
Probing Neuropeptide Volume Transmission In Vivo by Simultaneous Near-Infrared Light-Triggered Release and Optical Sensing.Angew Chem Int Ed Engl. 2022 Aug 22;61(34):e202206122. doi: 10.1002/anie.202206122. Epub 2022 Jul 8. Angew Chem Int Ed Engl. 2022. PMID: 35723610 Free PMC article.
References
-
- Romo R, and Schultz W. (1990). Dopamine neurons of the monkey midbrain: contingencies of responses to active touch during self-initiated arm movements. J. Neurophysiol. 63, 592–606. - PubMed
-
- Phillips PEM, Stuber GD, Heien MLAV, Wightman RM, and Carelli RM (2003). Subsecond dopamine release promotes cocaine seeking. Nature 422, 614–618. - PubMed
