

A Structural Study of the Cytoplasmic Chaperone Effect of 14-3-3 Proteins on Ataxin-1
- PMID: 34302818
- PMCID: PMC8505757
- DOI: 10.1016/j.jmb.2021.167174
A Structural Study of the Cytoplasmic Chaperone Effect of 14-3-3 Proteins on Ataxin-1
Abstract
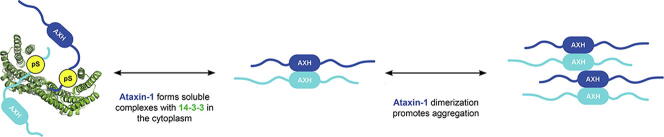
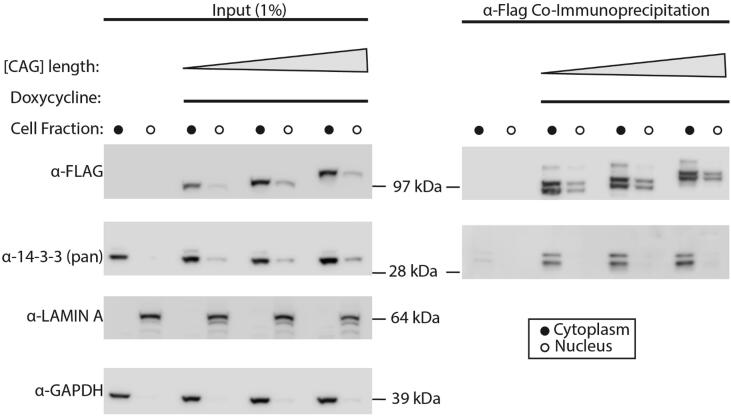
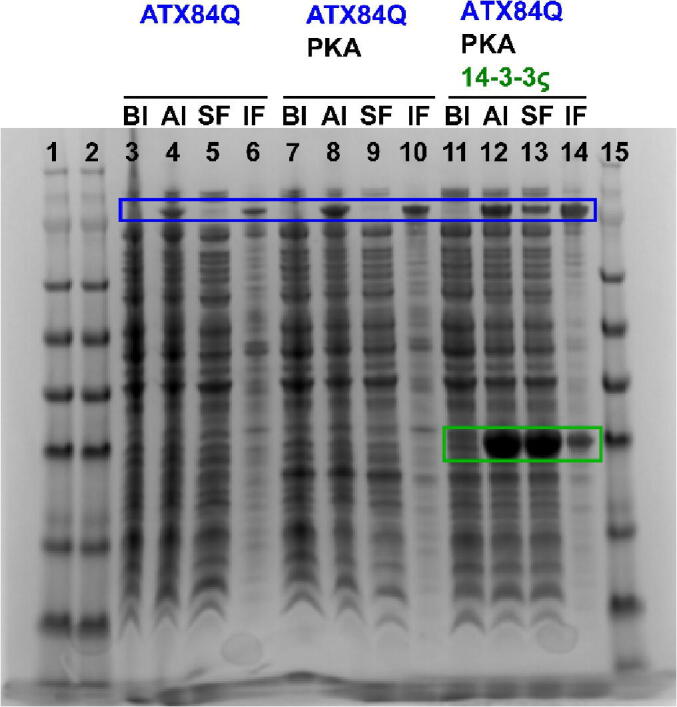
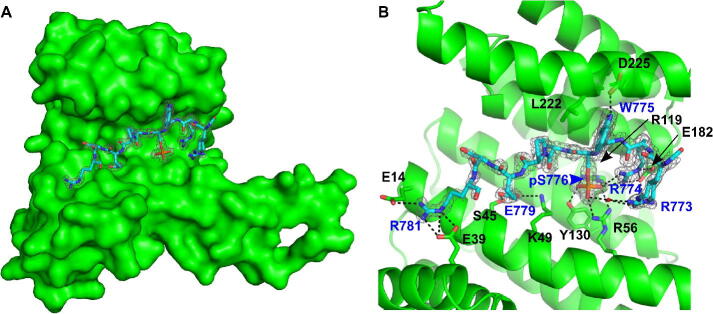
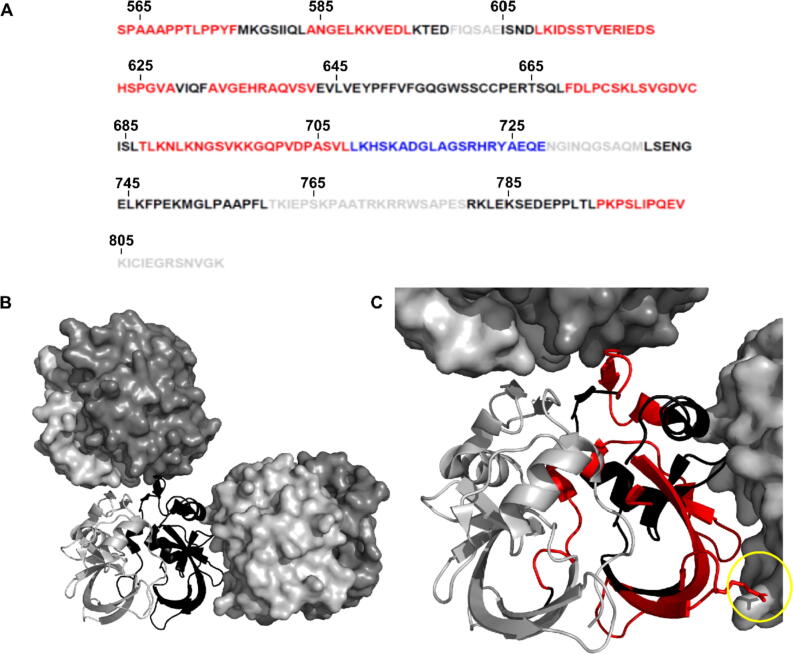
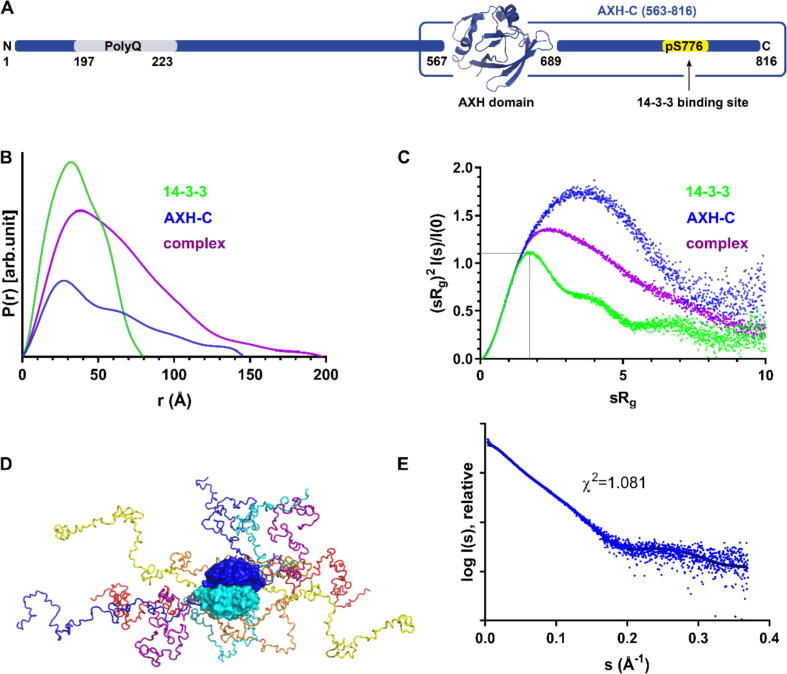
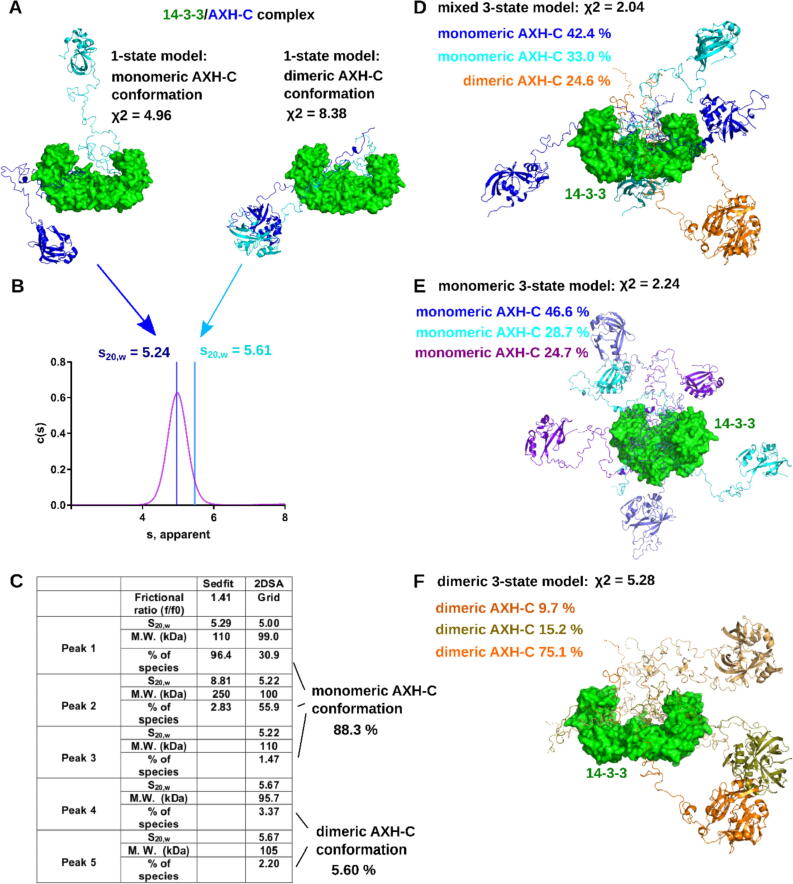
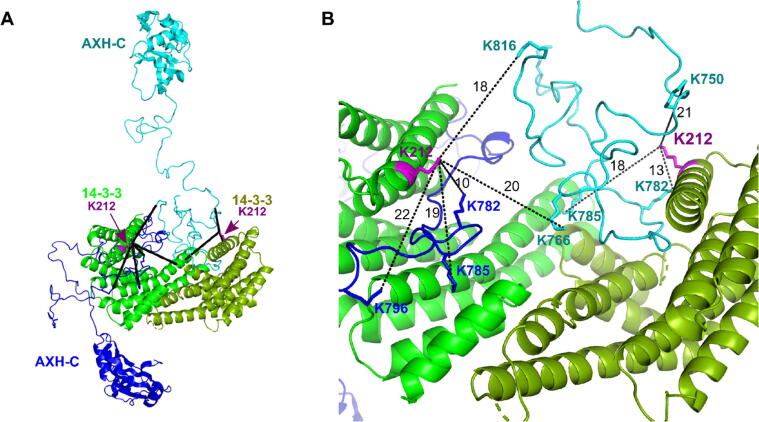
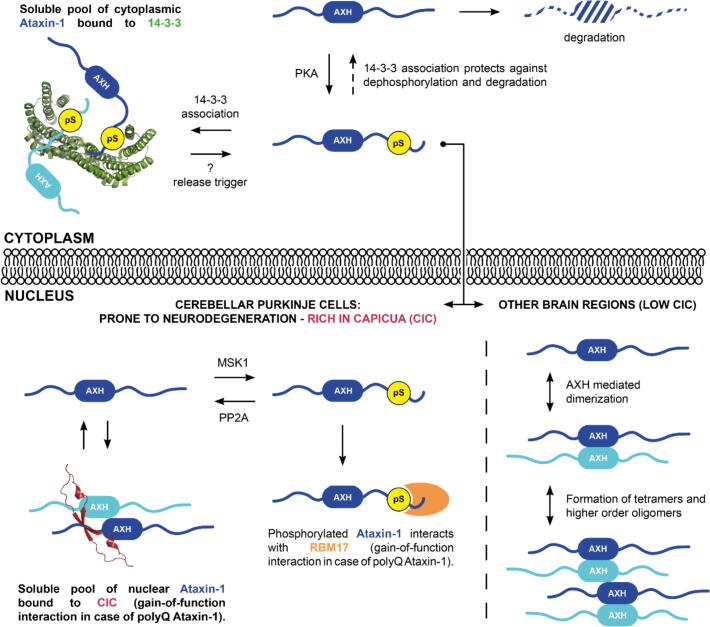
Expansion of the polyglutamine tract in the N terminus of Ataxin-1 is the main cause of the neurodegenerative disease, spinocerebellar ataxia type 1 (SCA1). However, the C-terminal part of the protein - including its AXH domain and a phosphorylation on residue serine 776 - also plays a crucial role in disease development. This phosphorylation event is known to be crucial for the interaction of Ataxin-1 with the 14-3-3 adaptor proteins and has been shown to indirectly contribute to Ataxin-1 stability. Here we show that 14-3-3 also has a direct anti-aggregation or "chaperone" effect on Ataxin-1. Furthermore, we provide structural and biophysical information revealing how phosphorylated S776 in the intrinsically disordered C terminus of Ataxin-1 mediates the cytoplasmic interaction with 14-3-3 proteins. Based on these findings, we propose that 14-3-3 exerts the observed chaperone effect by interfering with Ataxin-1 dimerization through its AXH domain, reducing further self-association. The chaperone effect is particularly important in the context of SCA1, as it was previously shown that a soluble form of mutant Ataxin-1 is the major driver of pathology.
Keywords: HDX-MS; SAXS; crystal structure; neurodegeneration; protein aggregation.
Copyright © 2021 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Opal P., Ashizawa T. In: GeneReviews((R)) Adam M.P., Ardinger H.H., Pagon R.A., Wallace S.E., Bean L.J.H., Stephens K., editors. University of Washington; Seattle (WA): 1993. Spinocerebellar Ataxia Type 1. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
