

Cell cycle on the crossroad of tumorigenesis and cancer therapy
- PMID: 34304958
- PMCID: PMC8688170
- DOI: 10.1016/j.tcb.2021.07.001
Cell cycle on the crossroad of tumorigenesis and cancer therapy
Abstract
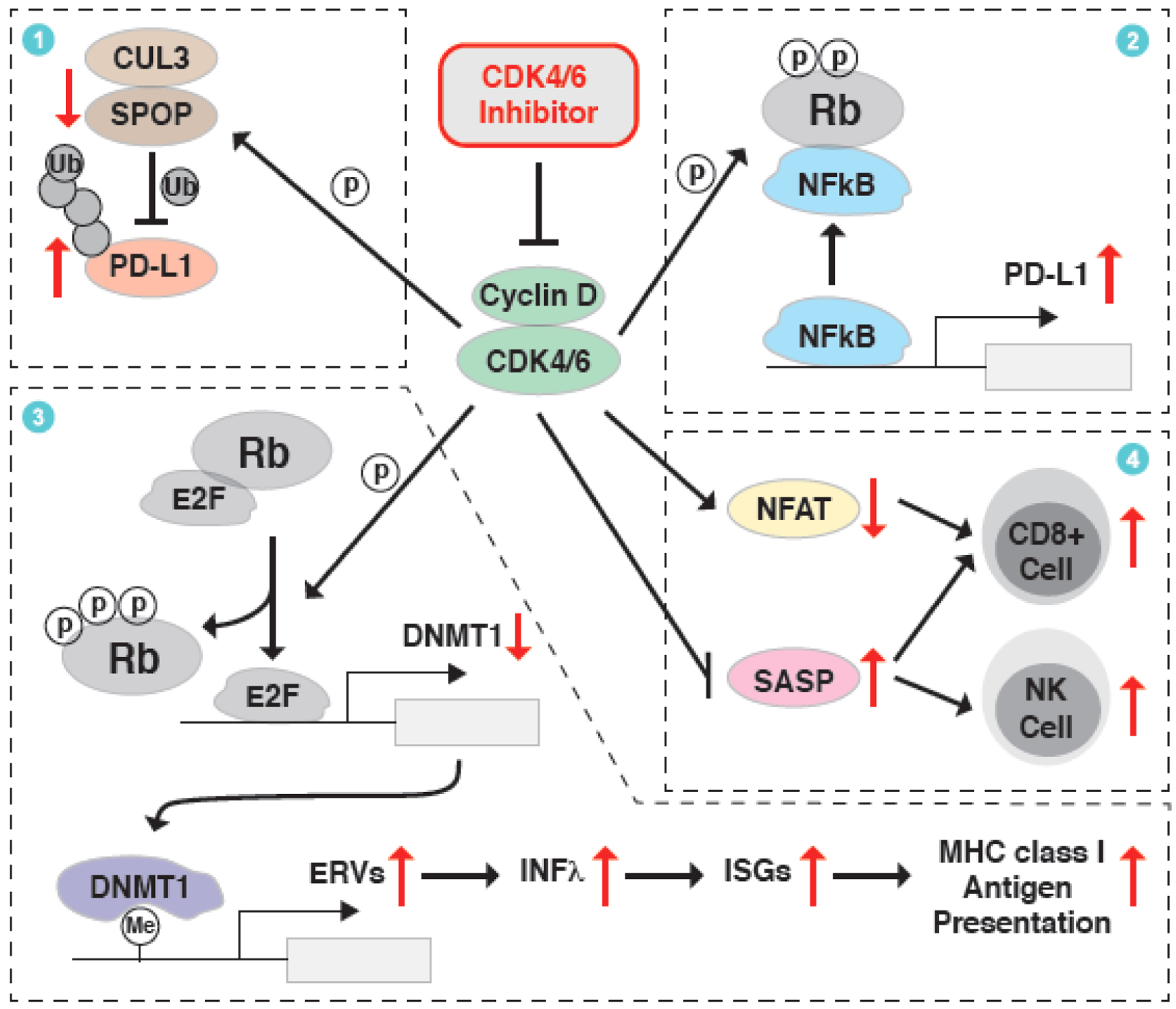
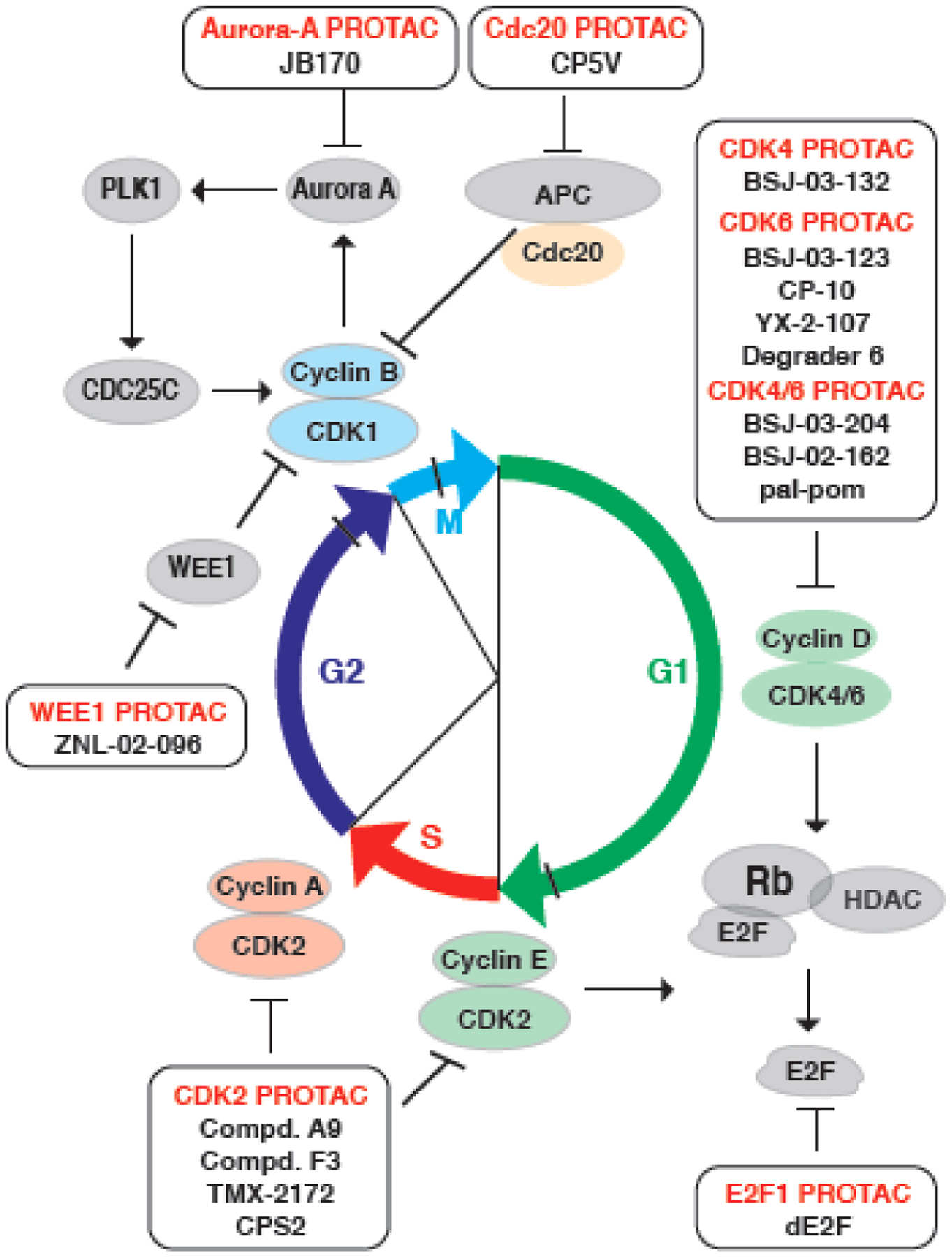
Aberrancy in cell cycle progression is one of the fundamental mechanisms underlying tumorigenesis, making regulators of the cell cycle machinery rational anticancer therapeutic targets. A growing body of evidence indicates that the cell cycle regulatory pathway integrates into other hallmarks of cancer, including metabolism remodeling and immune escape. Thus, therapies against cell cycle machinery components can not only repress the division of cancer cells, but also reverse cancer metabolism and restore cancer immune surveillance. Besides the ongoing effects on the development of small molecule inhibitors (SMIs) of the cell cycle machinery, proteolysis targeting chimeras (PROTACs) have recently been used to target these oncogenic proteins related to cell cycle progression. Here, we discuss the rationale of cell cycle targeting therapies, particularly PROTACs, to more efficiently retard tumorigenesis.
Keywords: PROTAC; cancer; cancer immune; cell cycle; degradation; metabolism.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests W.W. is a cofounder of and consultant for ReKindle Therapeutics. The other authors have no interests to declare.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
