

Exact Probability Landscapes of Stochastic Phenotype Switching in Feed-Forward Loops: Phase Diagrams of Multimodality
- PMID: 34306004
- PMCID: PMC8297706
- DOI: 10.3389/fgene.2021.645640
Exact Probability Landscapes of Stochastic Phenotype Switching in Feed-Forward Loops: Phase Diagrams of Multimodality
Abstract
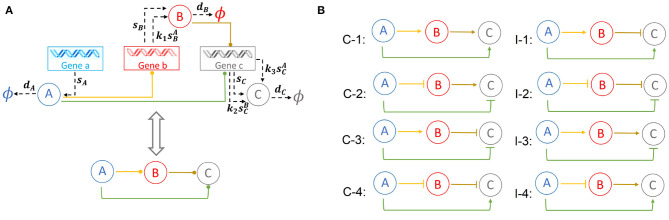
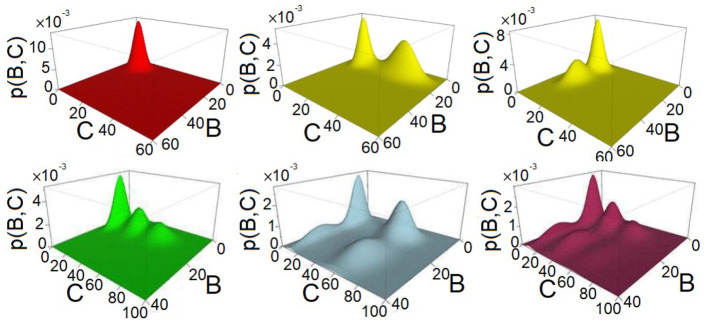
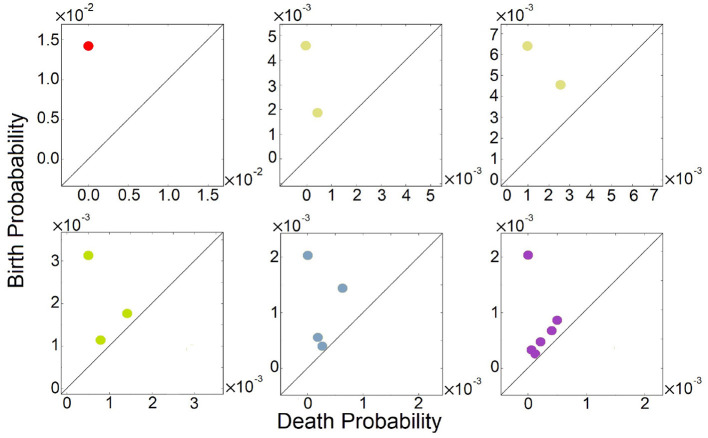
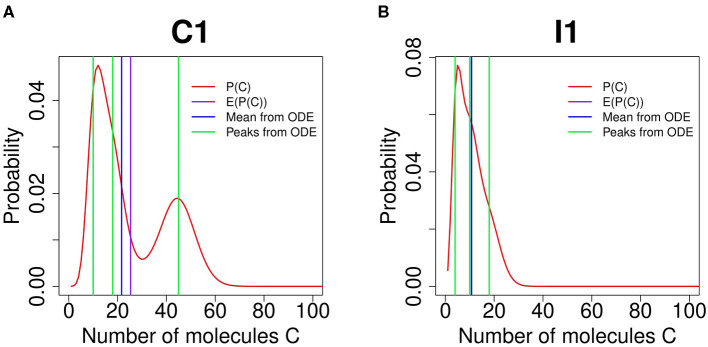
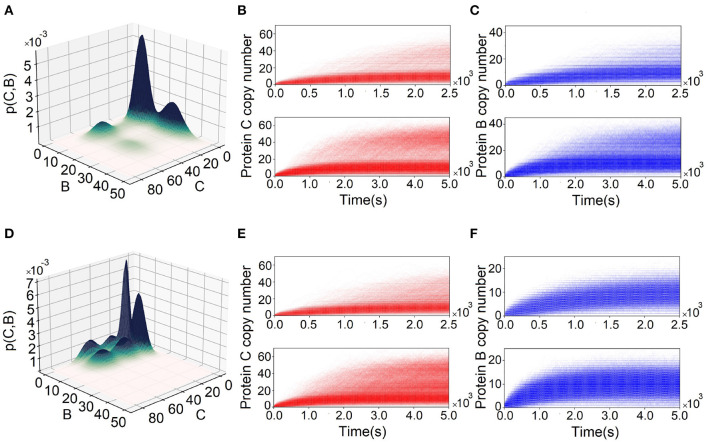
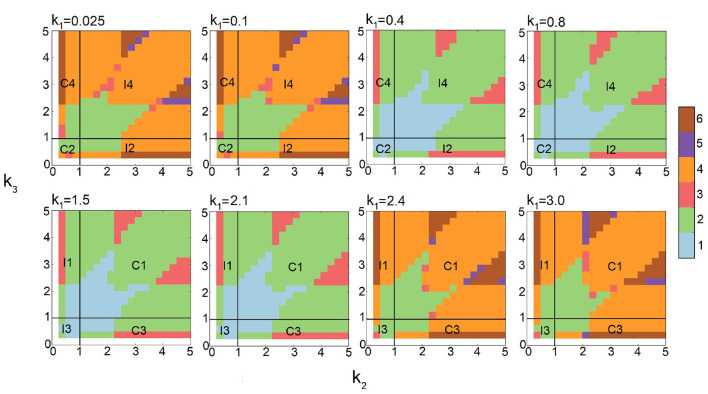
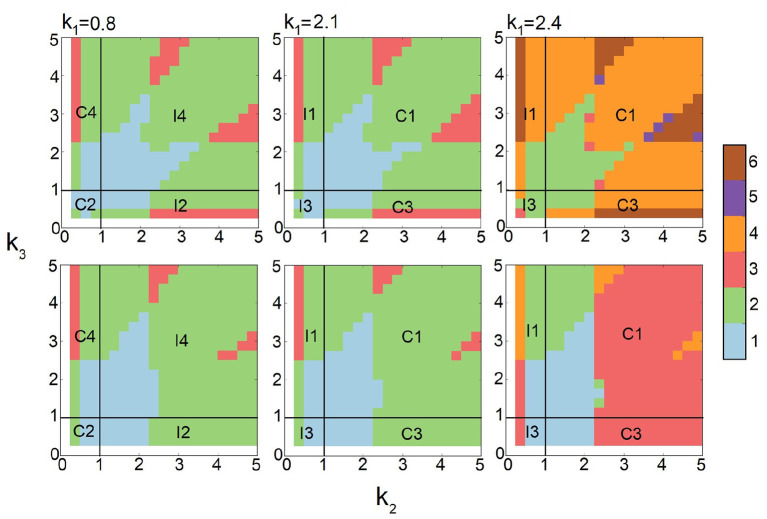
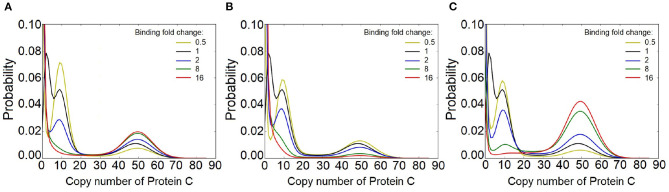
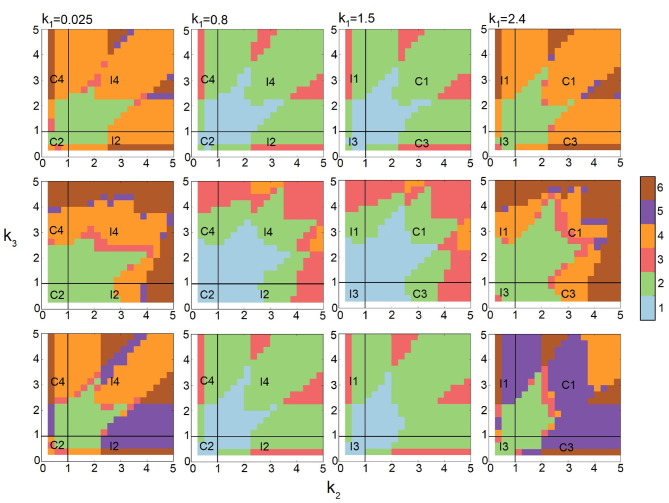
Feed-forward loops (FFLs) are among the most ubiquitously found motifs of reaction networks in nature. However, little is known about their stochastic behavior and the variety of network phenotypes they can exhibit. In this study, we provide full characterizations of the properties of stochastic multimodality of FFLs, and how switching between different network phenotypes are controlled. We have computed the exact steady-state probability landscapes of all eight types of coherent and incoherent FFLs using the finite-butter Accurate Chemical Master Equation (ACME) algorithm, and quantified the exact topological features of their high-dimensional probability landscapes using persistent homology. Through analysis of the degree of multimodality for each of a set of 10,812 probability landscapes, where each landscape resides over 105-106 microstates, we have constructed comprehensive phase diagrams of all relevant behavior of FFL multimodality over broad ranges of input and regulation intensities, as well as different regimes of promoter binding dynamics. In addition, we have quantified the topological sensitivity of the multimodality of the landscapes to regulation intensities. Our results show that with slow binding and unbinding dynamics of transcription factor to promoter, FFLs exhibit strong stochastic behavior that is very different from what would be inferred from deterministic models. In addition, input intensity play major roles in the phenotypes of FFLs: At weak input intensity, FFL exhibit monomodality, but strong input intensity may result in up to 6 stable phenotypes. Furthermore, we found that gene duplication can enlarge stable regions of specific multimodalities and enrich the phenotypic diversity of FFL networks, providing means for cells toward better adaptation to changing environment. Our results are directly applicable to analysis of behavior of FFLs in biological processes such as stem cell differentiation and for design of synthetic networks when certain phenotypic behavior is desired.
Keywords: ACME algorithm; feed forward loop; gene regulatory network; network motif; persistent homology; stochastic reaction network; systems biology.
Copyright © 2021 Terebus, Manuchehrfar, Cao and Liang.
Conflict of interest statement
YC was employed by company Merck & Co., Inc., Kenilworth, NJ, United States. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Sensitivities of Regulation Intensities in Feed-Forward Loops with Multistability.Annu Int Conf IEEE Eng Med Biol Soc. 2019 Jul;2019:1969-1972. doi: 10.1109/EMBC.2019.8856532. Annu Int Conf IEEE Eng Med Biol Soc. 2019. PMID: 31946285 Free PMC article.
-
Dynamics of two feed forward genetic motifs in the presence of molecular noise.Biosystems. 2024 Dec;246:105352. doi: 10.1016/j.biosystems.2024.105352. Epub 2024 Oct 20. Biosystems. 2024. PMID: 39433119
-
Noise characteristics of feed forward loops.Phys Biol. 2005 Mar;2(1):36-45. doi: 10.1088/1478-3967/2/1/005. Phys Biol. 2005. PMID: 16204855
-
Transcription factor and microRNA co-regulatory loops: important regulatory motifs in biological processes and diseases.Brief Bioinform. 2015 Jan;16(1):45-58. doi: 10.1093/bib/bbt085. Epub 2013 Dec 4. Brief Bioinform. 2015. PMID: 24307685 Review.
-
Processes on the emergent landscapes of biochemical reaction networks and heterogeneous cell population dynamics: differentiation in living matters.J R Soc Interface. 2017 May;14(130):20170097. doi: 10.1098/rsif.2017.0097. J R Soc Interface. 2017. PMID: 28490602 Free PMC article. Review.
Cited by
-
Identifying Transient Cells During Reprogramming via Persistent Homology.Annu Int Conf IEEE Eng Med Biol Soc. 2022 Jul;2022:2920-2923. doi: 10.1109/EMBC48229.2022.9871358. Annu Int Conf IEEE Eng Med Biol Soc. 2022. PMID: 36085927 Free PMC article.
-
Allelic correlation is a marker of trade-offs between barriers to transmission of expression variability and signal responsiveness in genetic networks.Cell Syst. 2022 Dec 21;13(12):1016-1032.e6. doi: 10.1016/j.cels.2022.10.008. Epub 2022 Nov 29. Cell Syst. 2022. PMID: 36450286 Free PMC article.
-
Network design principle for robust oscillatory behaviors with respect to biological noise.Elife. 2022 Sep 20;11:e76188. doi: 10.7554/eLife.76188. Elife. 2022. PMID: 36125857 Free PMC article.
References
-
- Alon U. (2006). An Introduction to Systems Biology: Design Principles of Biological Circuits. New York, NY: CRC Press. 10.1201/9781420011432 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources