

Chronic methamphetamine uncovers a circadian rhythm in multiple-unit neural activity in the dorsal striatum which is independent of the suprachiasmatic nucleus
- PMID: 34307964
- PMCID: PMC8258683
- DOI: 10.1016/j.nbscr.2021.100070
Chronic methamphetamine uncovers a circadian rhythm in multiple-unit neural activity in the dorsal striatum which is independent of the suprachiasmatic nucleus
Abstract
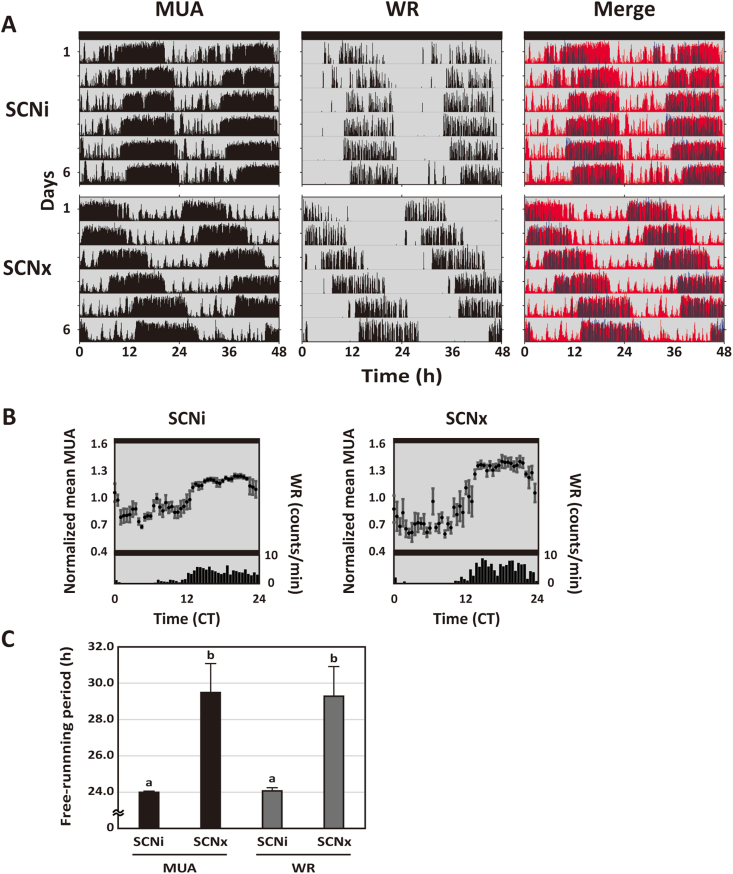
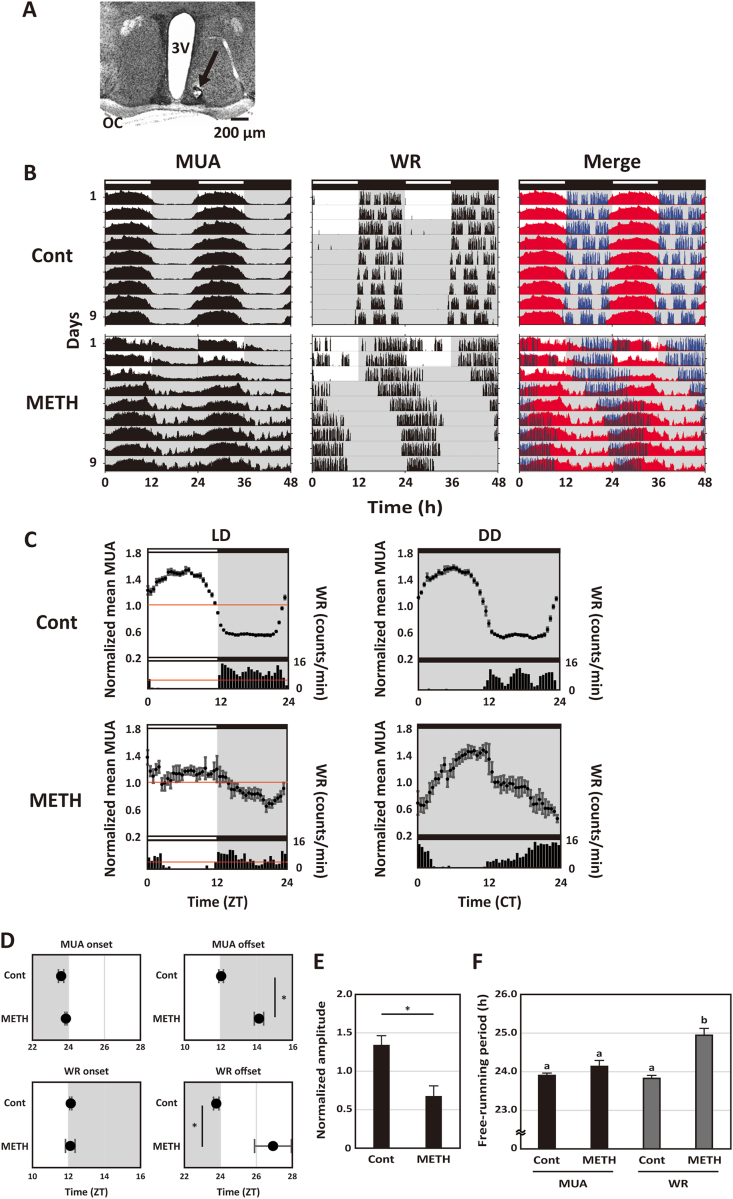
The dorsal striatum forms part of the basal ganglia circuit that is a major regulator of voluntary motor behavior. Dysfunction in this circuit is a critical factor in the pathology of neurological (Parkinson's and Huntington's disease) as well as psychiatric disorders. In this study, we employed in vivo real-time monitoring of multiple unit neural activity (MUA) in the dorsal striatum of freely moving mice. We demonstrate that the striatum exhibits robust diurnal and circadian rhythms in MUA that peak in the night. These rhythms are dependent upon the central circadian clock located in the suprachiasmatic nucleus (SCN) as lesions of this structure caused the loss of rhythmicity measured in the striatum. Nonetheless, chronic treatment of methamphetamine (METH) makes circadian rhythms appear in MUA recorded from the striatum of SCN-lesioned mice. These data demonstrate that the physiological properties of neurons in the dorsal striatum are regulated by the circadian system and that METH drives circadian rhythms in striatal physiology in the absence of the SCN. The finding of SCN-driven circadian rhythms in striatal physiology has important implications for an understanding of the temporal regulation of motor control as well as revealing how disease processes may disrupt this regulation.
Keywords: Circadian rhythm; Dopamine; Methamphetamine; Multiple unit neural activity; Striatum; Suprachiasmatic nucleus; Wheel running activity; constant dark, (DD); dopamine transporter, (DAT); dopamine, (DA); light-dark, (LD); methamphetamine, (METH); multiple unit neural activity, (MUA); suprachiasmatic nucleus, (SCN); wheel running, (WR).
© 2021 The Authors.
Conflict of interest statement
None.
Figures

References
LinkOut - more resources
Full Text Sources
Research Materials
