

Development of an Acrylamide-Based Inhibitor of Protein S-Acylation
- PMID: 34309372
- PMCID: PMC8590885
- DOI: 10.1021/acschembio.1c00405
Development of an Acrylamide-Based Inhibitor of Protein S-Acylation
Abstract
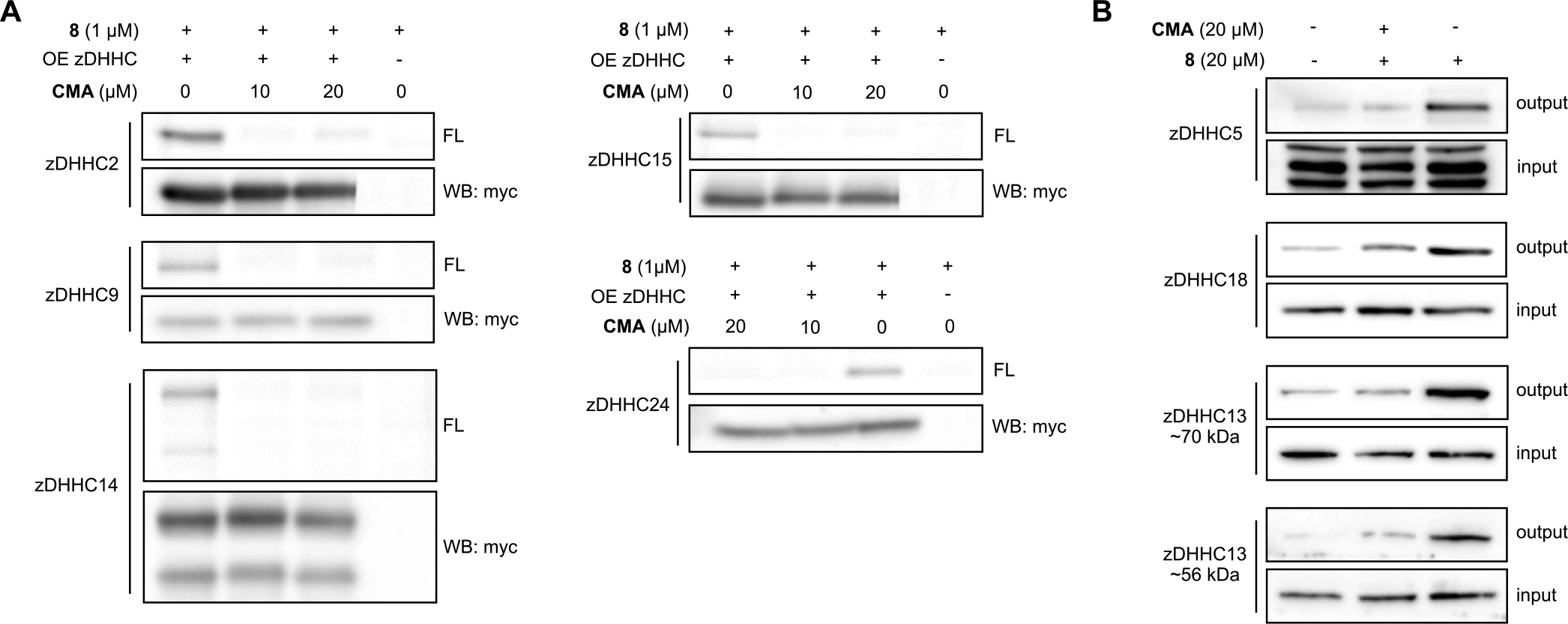
Protein S-acylation is a dynamic lipid post-translational modification that can modulate the localization and activity of target proteins. In humans, the installation of the lipid onto target proteins is catalyzed by a family of 23 Asp-His-His-Cys domain-containing protein acyltransferases (DHHC-PATs). DHHCs are increasingly recognized as critical players in cellular signaling events and in human disease. However, progress elucidating the functions and mechanisms of DHHC "writers" has been hampered by a lack of chemical tools to perturb their activity in live cells. Herein, we report the synthesis and characterization of cyano-myracrylamide (CMA), a broad-spectrum DHHC family inhibitor with similar potency to 2-bromopalmitate (2BP), the most commonly used DHHC inhibitor in the field. Possessing an acrylamide warhead instead of 2BP's α-halo fatty acid, CMA inhibits DHHC family proteins in cellulo while demonstrating decreased toxicity and avoiding inhibition of the S-acylation eraser enzymes, two of the major weaknesses of 2BP. Our studies show that CMA engages with DHHC family proteins in cells, inhibits protein S-acylation, and disrupts DHHC-regulated cellular events. CMA represents an improved chemical scaffold for untangling the complexities of DHHC-mediated cell signaling by protein S-acylation.
Conflict of interest statement
The authors declare the following competing financial interest(s): R.S.K. and B.C.D. have a patent on the DPP technology used in this work.
Figures




References
-
- Lanyon-Hogg T, Faronato M, Serwa RA, and Tate EW (2017) Dynamic Protein Acylation: New Substrates, Mechanisms, and Drug Targets. Trends Biochem. Sci. 42, 566–581. - PubMed
-
- Linder ME, and Deschenes RJ (2007) Palmitoylation: policing protein stability and traffic. Nat. Rev. Mol. Cell Biol. 8, 74–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
