

Autoregulatory control of microtubule binding in doublecortin-like kinase 1
- PMID: 34310279
- PMCID: PMC8352597
- DOI: 10.7554/eLife.60126
Autoregulatory control of microtubule binding in doublecortin-like kinase 1
Abstract
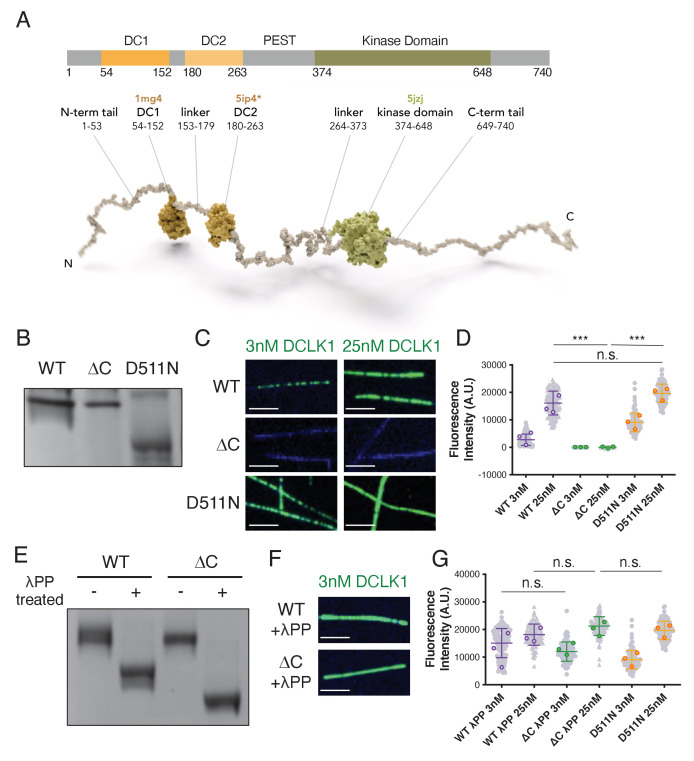
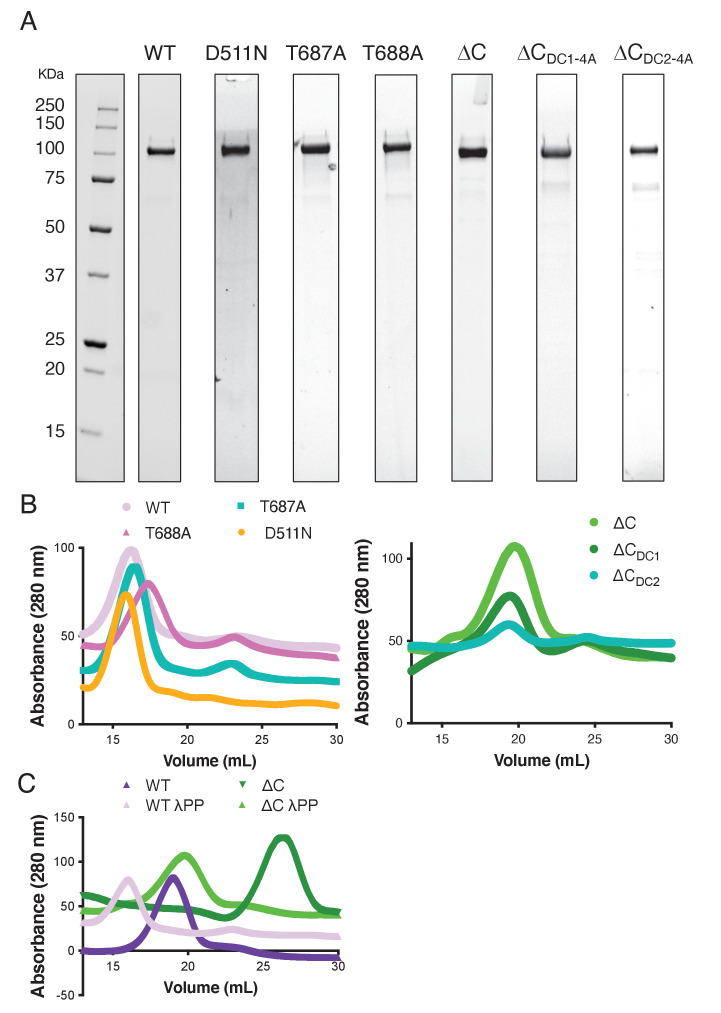
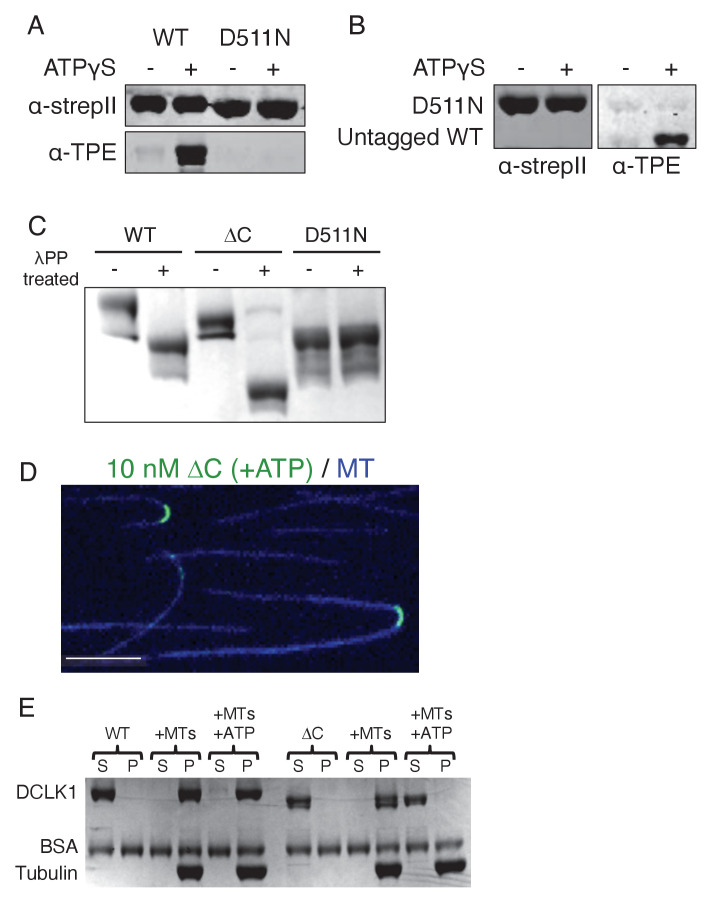
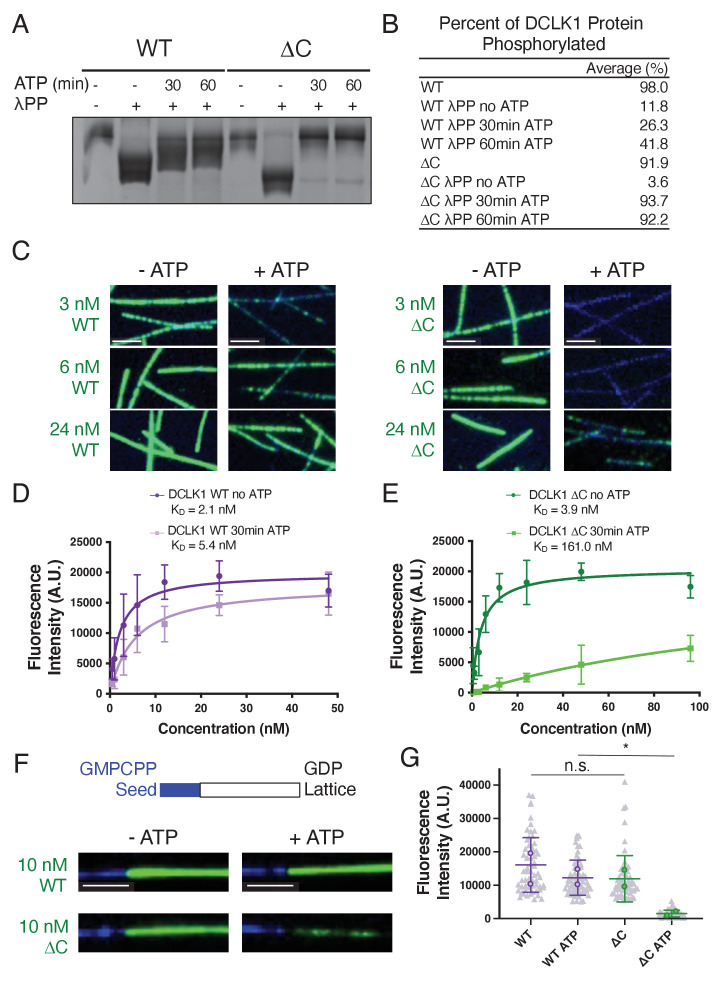
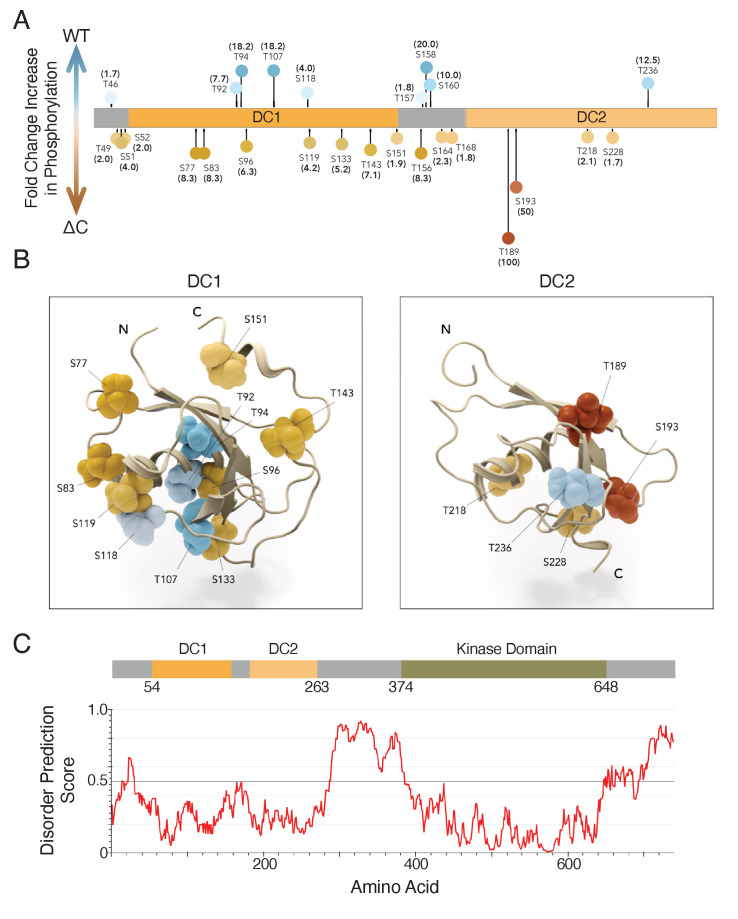
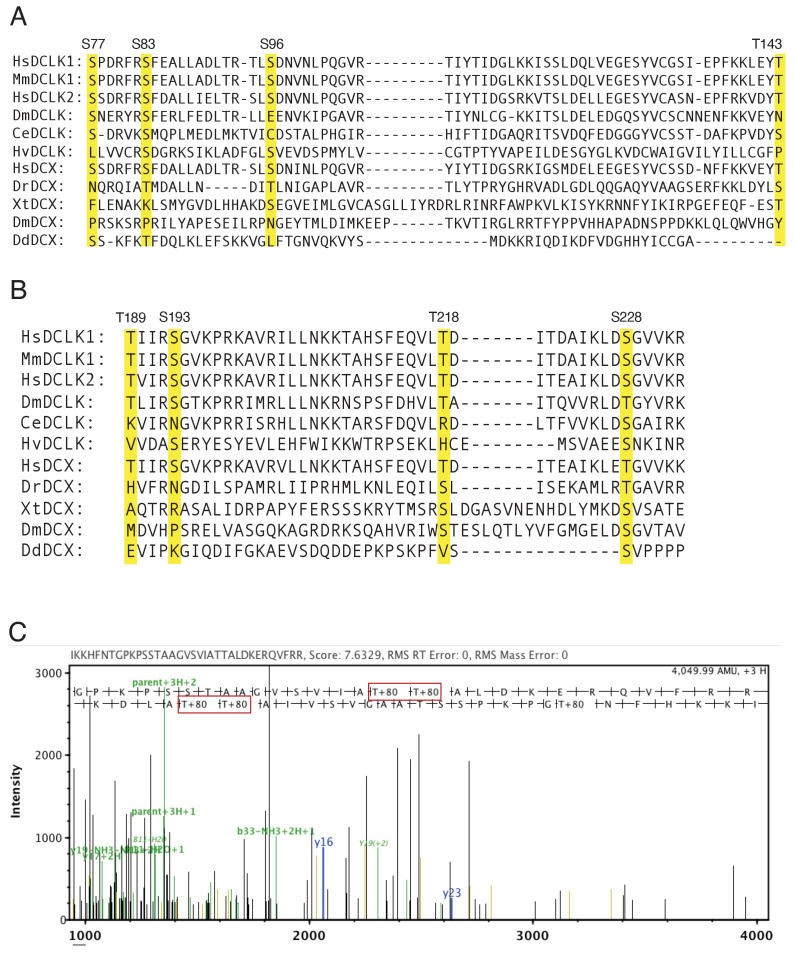
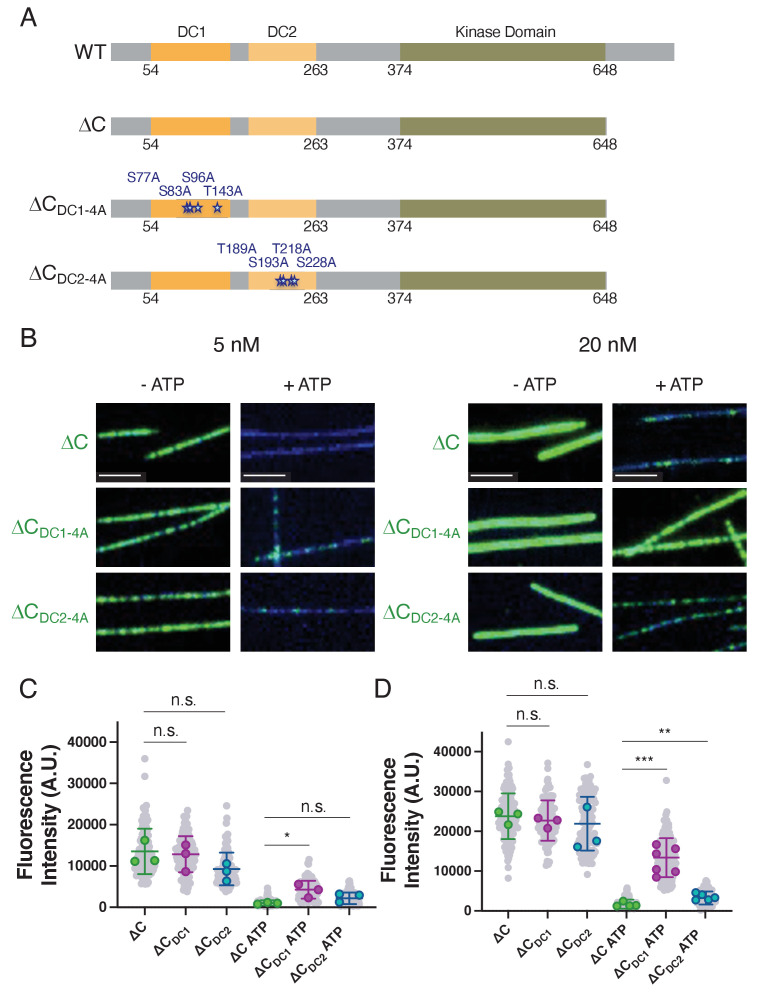
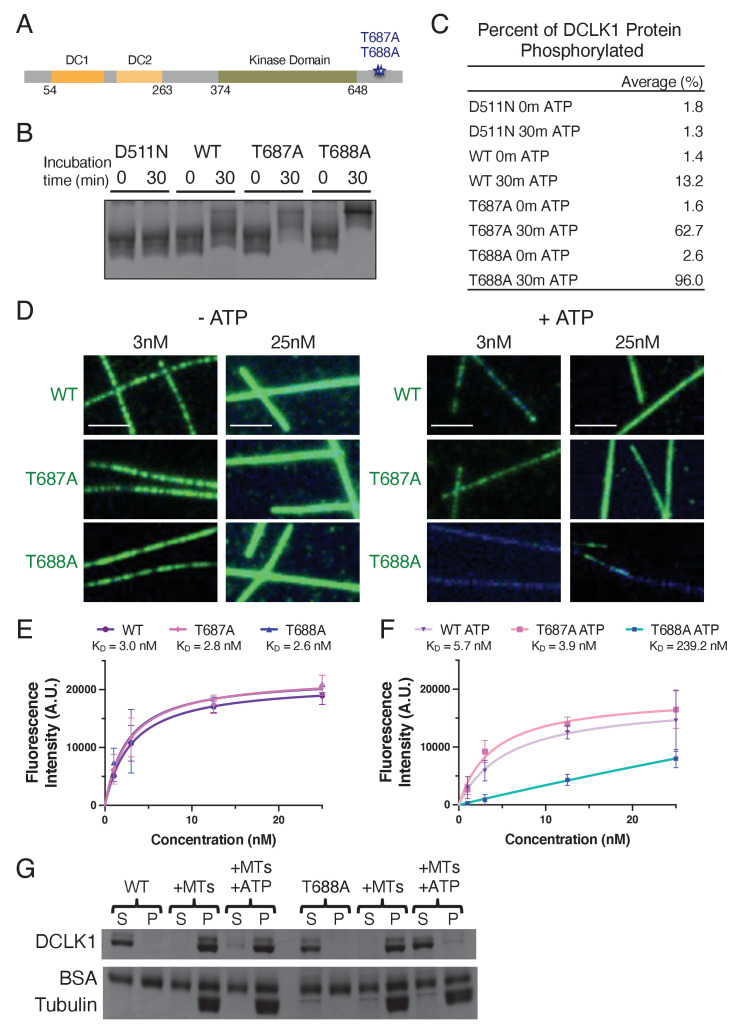
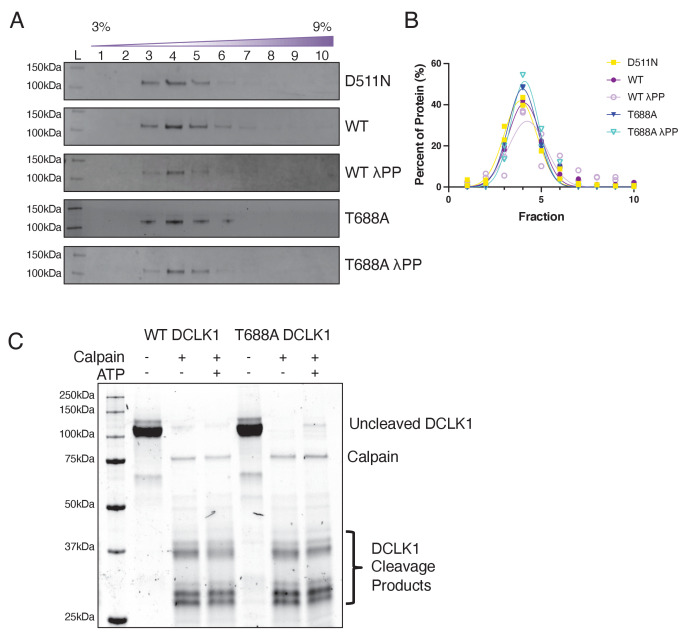
The microtubule-associated protein, doublecortin-like kinase 1 (DCLK1), is highly expressed in a range of cancers and is a prominent therapeutic target for kinase inhibitors. The physiological roles of DCLK1 kinase activity and how it is regulated remain elusive. Here, we analyze the role of mammalian DCLK1 kinase activity in regulating microtubule binding. We found that DCLK1 autophosphorylates a residue within its C-terminal tail to restrict its kinase activity and prevent aberrant hyperphosphorylation within its microtubule-binding domain. Removal of the C-terminal tail or mutation of this residue causes an increase in phosphorylation within the doublecortin domains, which abolishes microtubule binding. Therefore, autophosphorylation at specific sites within DCLK1 has diametric effects on the molecule's association with microtubules. Our results suggest a mechanism by which DCLK1 modulates its kinase activity to tune its microtubule-binding affinity. These results provide molecular insights for future therapeutic efforts related to DCLK1's role in cancer development and progression.
Keywords: MAP; autophosphorylation; biochemistry; cancer; cancer biology; chemical biology; doublecortin-like kinase 1; microtubule; microtubule-associated protein; mouse.
© 2021, Agulto et al.
Conflict of interest statement
RA, MR, TT, AR, AD, HB, JC, DN No competing interests declared, KO Reviewing editor, eLife
Figures
References
-
- Burger D, Stihle M, Sharma A, Di Lello P, Benz J, D'Arcy B, Debulpaep M, Fry D, Huber W, Kremer T, Laeremans T, Matile H, Ross A, Rufer AC, Schoch G, Steinmetz MO, Steyaert J, Rudolph MG, Thoma R, Ruf A. Crystal structures of the human doublecortin C- and N-terminal domains in complex with specific antibodies. Journal of Biological Chemistry. 2016;291:16292–16306. doi: 10.1074/jbc.M116.726547. - DOI - PMC - PubMed
