

Epigenetic scars of CD8+ T cell exhaustion persist after cure of chronic infection in humans
- PMID: 34312547
- PMCID: PMC8600539
- DOI: 10.1038/s41590-021-00979-1
Epigenetic scars of CD8+ T cell exhaustion persist after cure of chronic infection in humans
Abstract
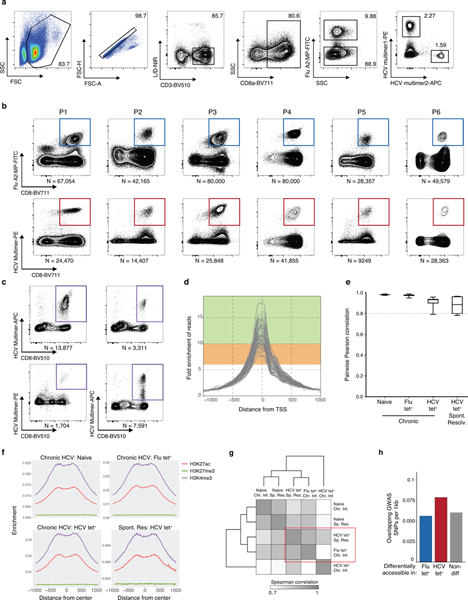
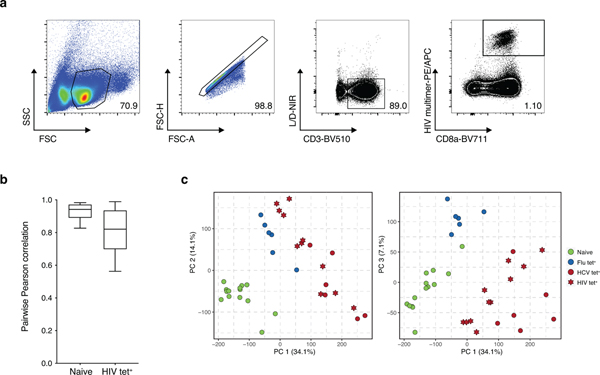
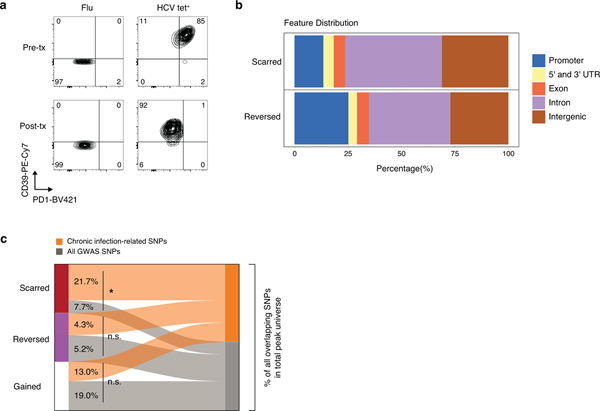
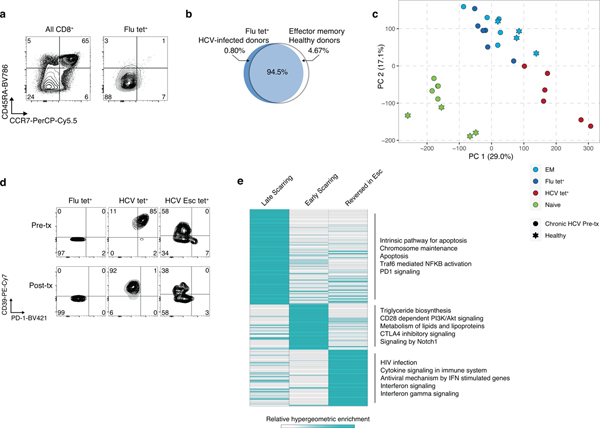
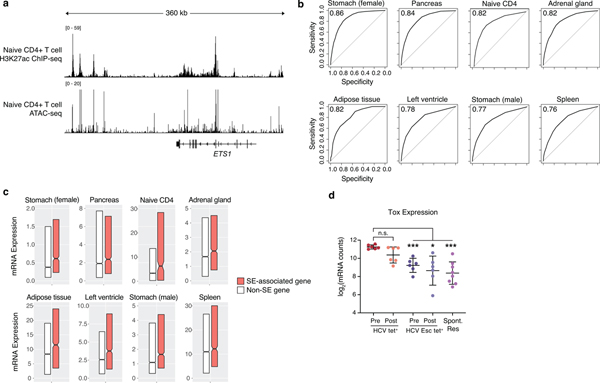
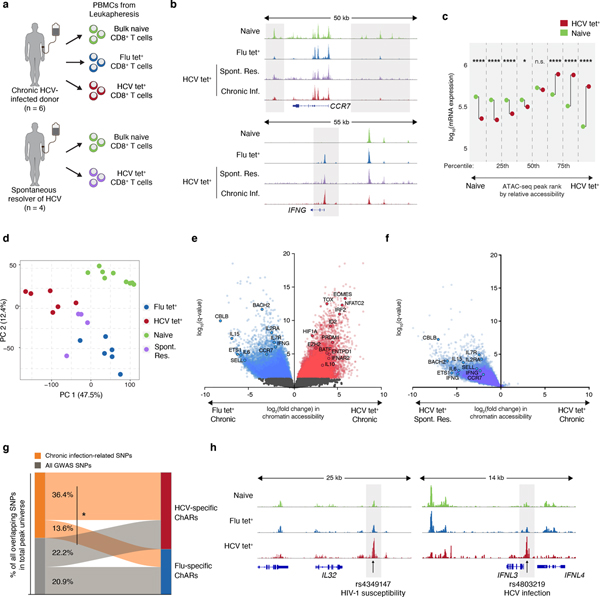
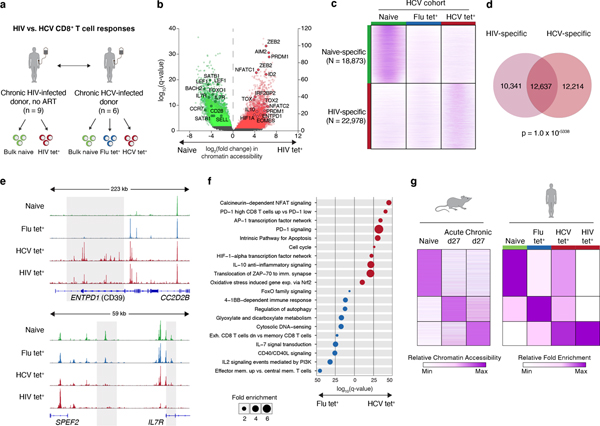
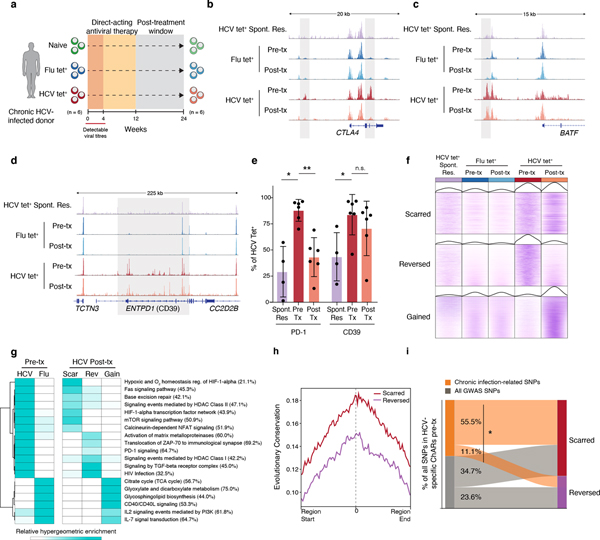
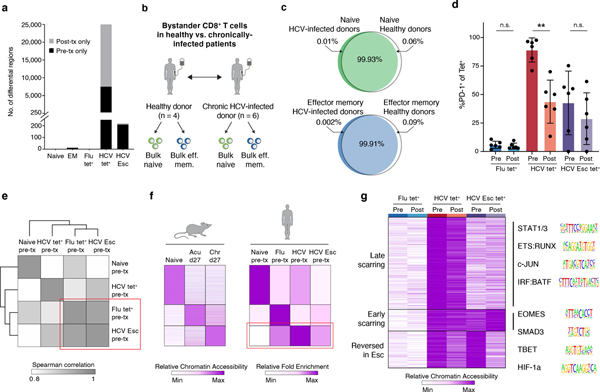
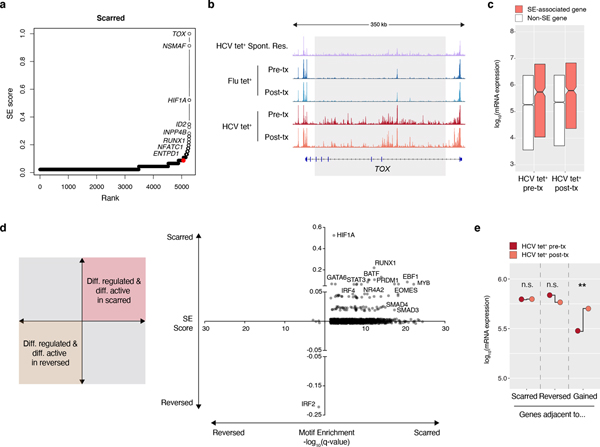
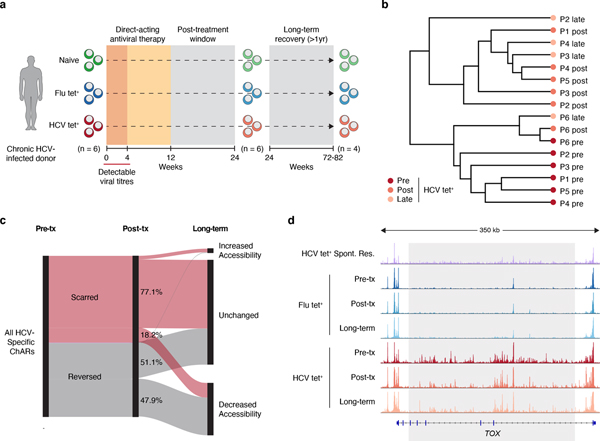
T cell exhaustion is an induced state of dysfunction that arises in response to chronic infection and cancer. Exhausted CD8+ T cells acquire a distinct epigenetic state, but it is not known whether that chromatin landscape is fixed or plastic following the resolution of a chronic infection. Here we show that the epigenetic state of exhaustion is largely irreversible, even after curative therapy. Analysis of chromatin accessibility in HCV- and HIV-specific responses identifies a core epigenetic program of exhaustion in CD8+ T cells, which undergoes only limited remodeling before and after resolution of infection. Moreover, canonical features of exhaustion, including super-enhancers near the genes TOX and HIF1A, remain 'epigenetically scarred.' T cell exhaustion is therefore a conserved epigenetic state that becomes fixed and persists independent of chronic antigen stimulation and inflammation. Therapeutic efforts to reverse T cell exhaustion may require new approaches that increase the epigenetic plasticity of exhausted T cells.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests
AbbVie sponsored the clinical trial (
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
