

ELAVL4, splicing, and glutamatergic dysfunction precede neuron loss in MAPT mutation cerebral organoids
- PMID: 34314701
- PMCID: PMC8635409
- DOI: 10.1016/j.cell.2021.07.003
ELAVL4, splicing, and glutamatergic dysfunction precede neuron loss in MAPT mutation cerebral organoids
Abstract
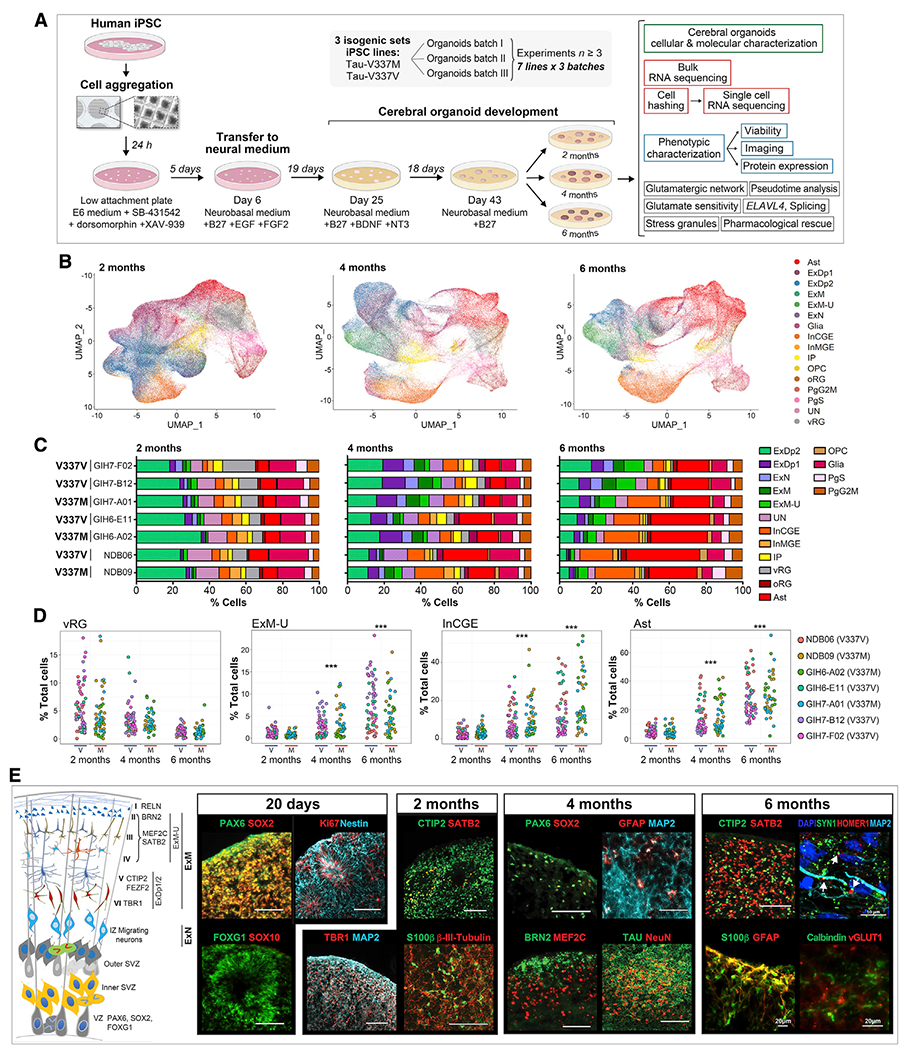
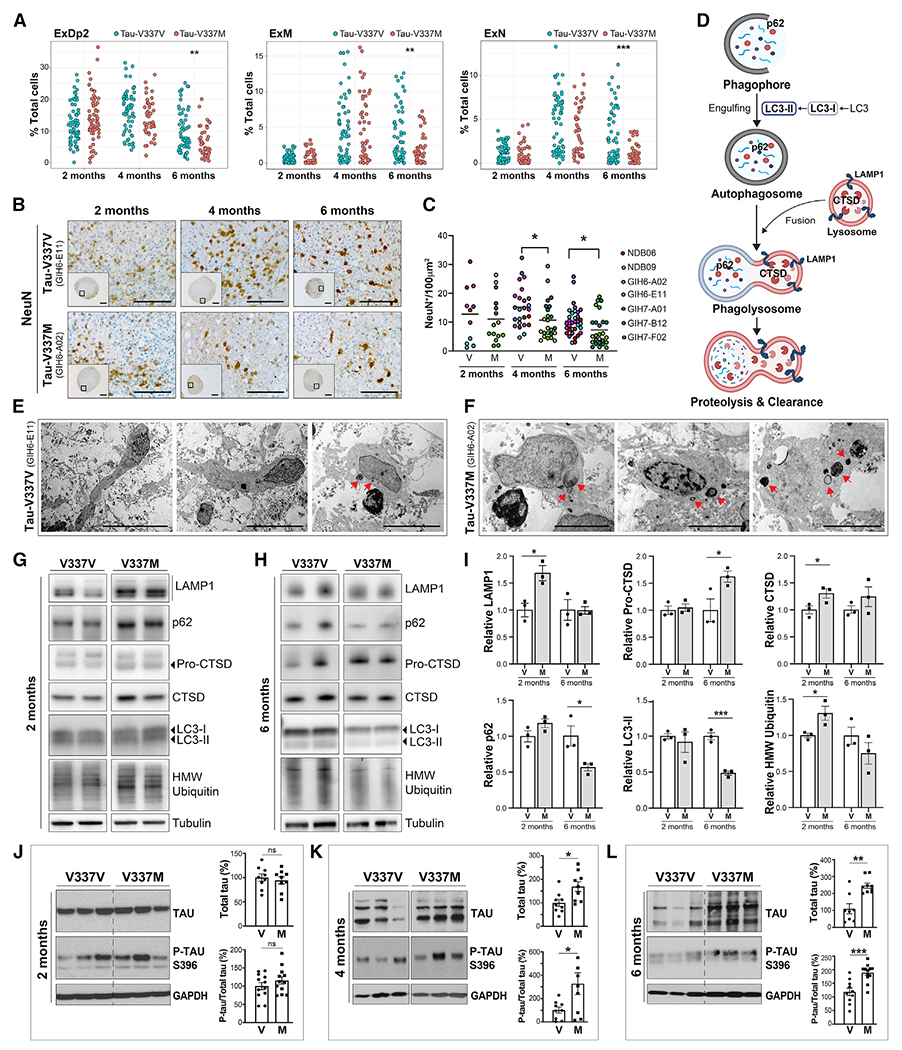
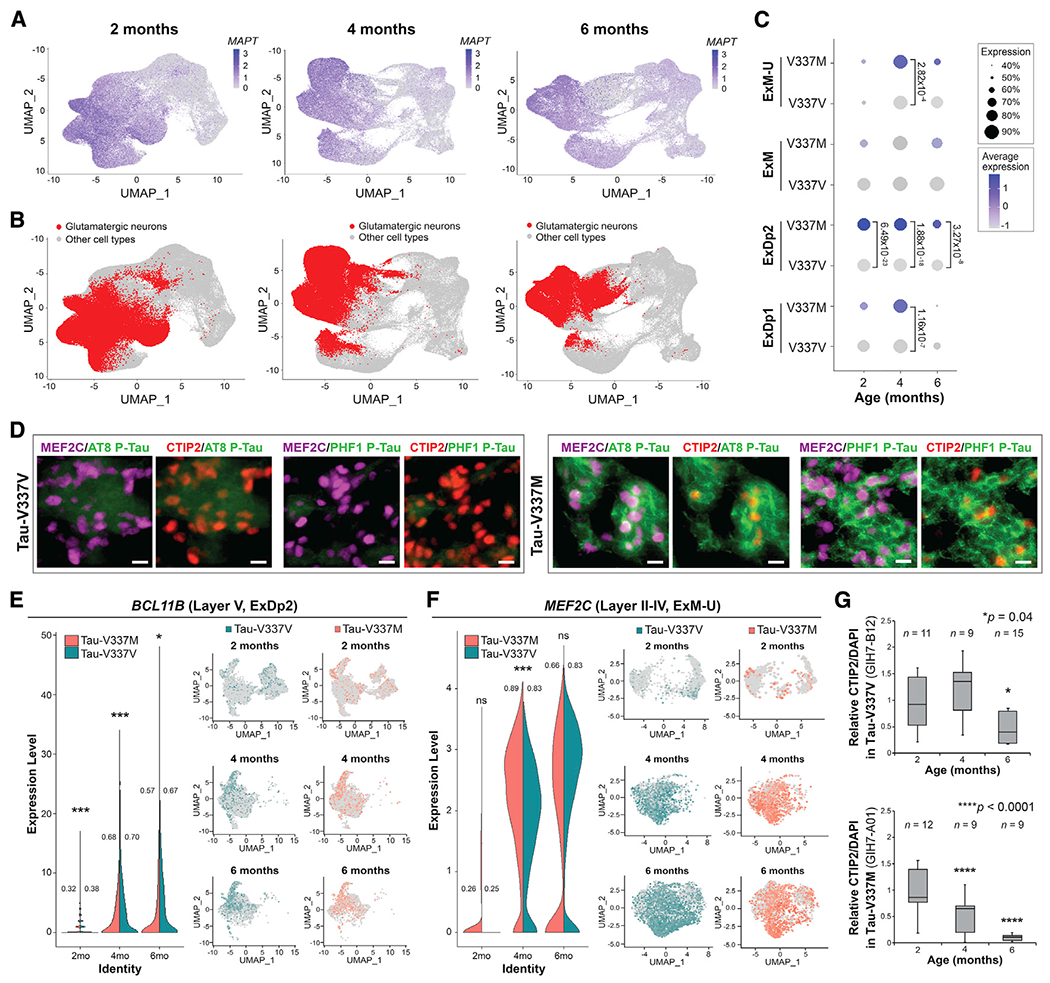
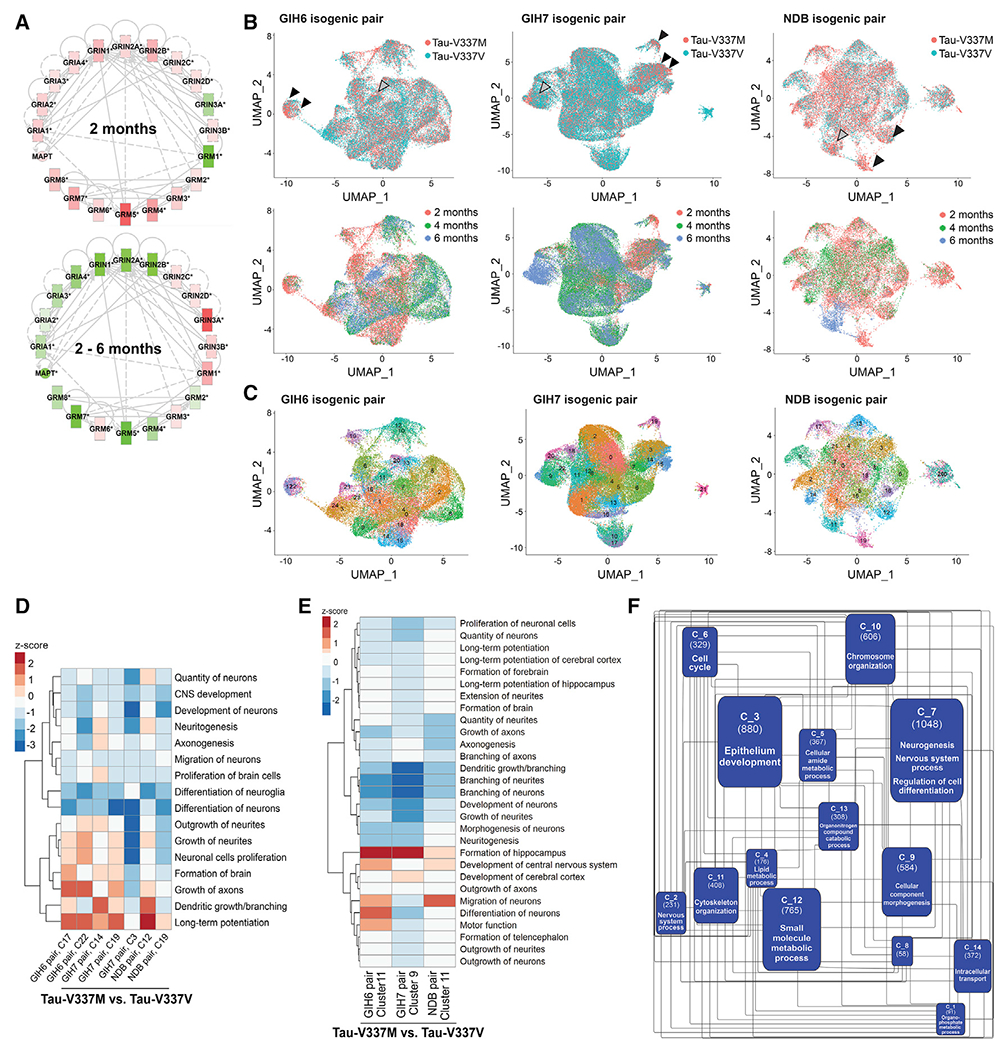
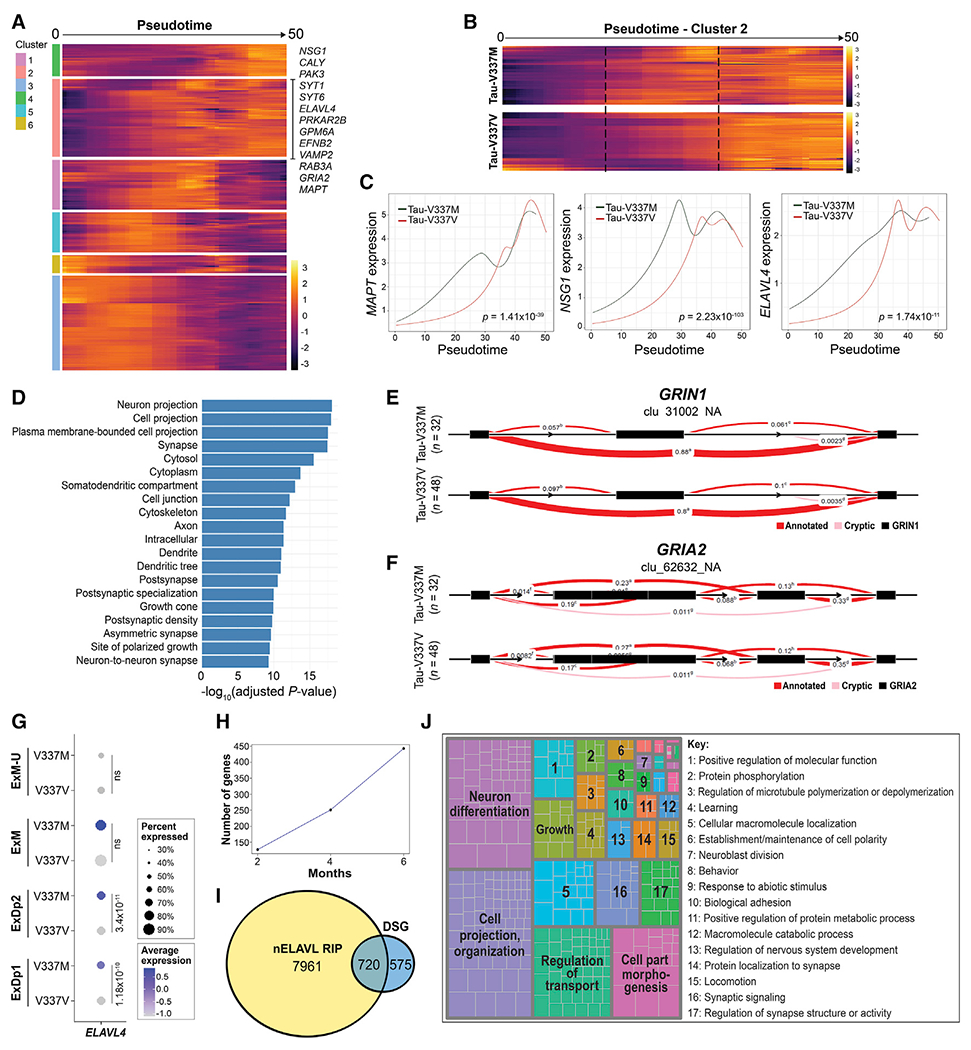
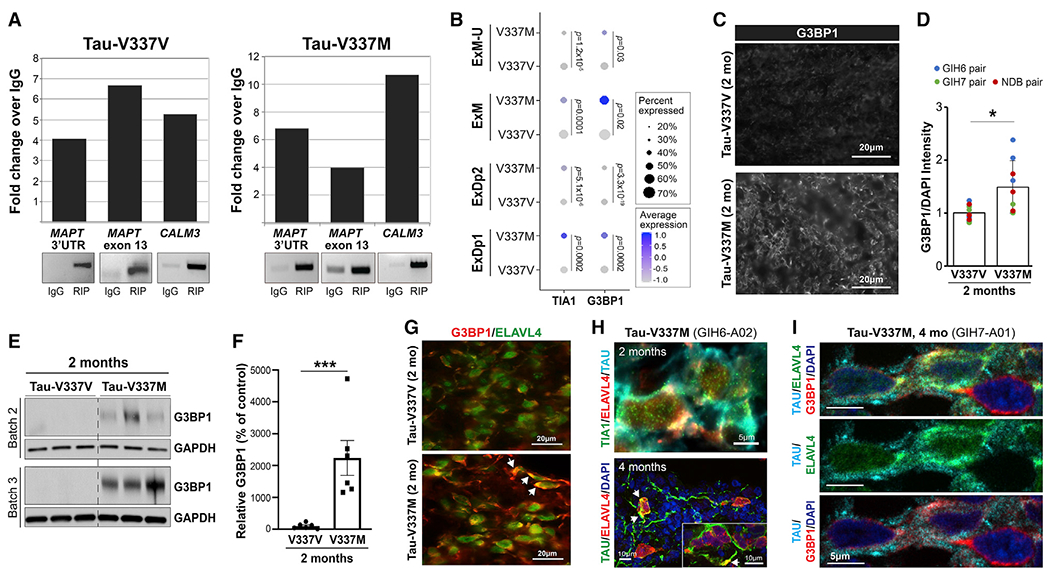
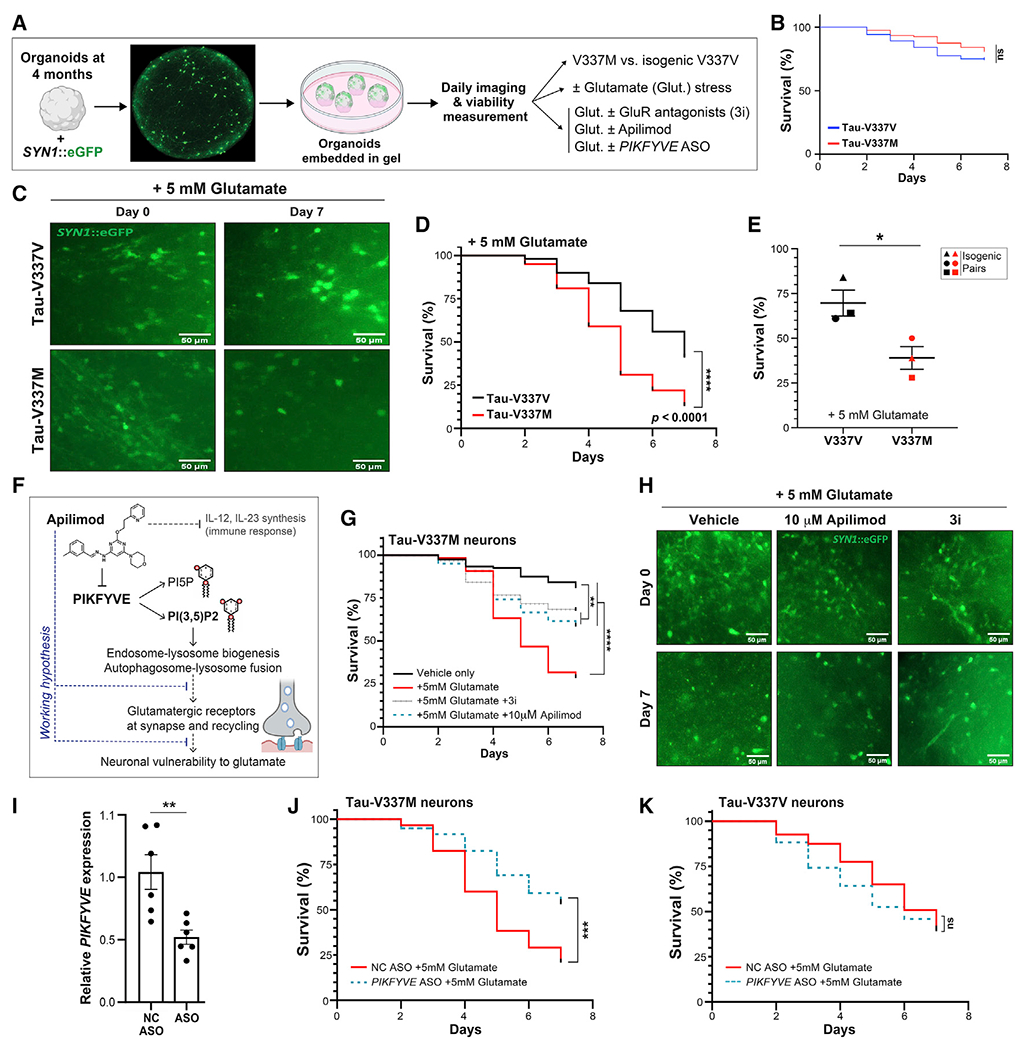
Frontotemporal dementia (FTD) because of MAPT mutation causes pathological accumulation of tau and glutamatergic cortical neuronal death by unknown mechanisms. We used human induced pluripotent stem cell (iPSC)-derived cerebral organoids expressing tau-V337M and isogenic corrected controls to discover early alterations because of the mutation that precede neurodegeneration. At 2 months, mutant organoids show upregulated expression of MAPT, glutamatergic signaling pathways, and regulators, including the RNA-binding protein ELAVL4, and increased stress granules. Over the following 4 months, mutant organoids accumulate splicing changes, disruption of autophagy function, and build-up of tau and P-tau-S396. By 6 months, tau-V337M organoids show specific loss of glutamatergic neurons as seen in individuals with FTD. Mutant neurons are susceptible to glutamate toxicity, which can be rescued pharmacologically by the PIKFYVE kinase inhibitor apilimod. Our results demonstrate a sequence of events that precede neurodegeneration, revealing molecular pathways associated with glutamate signaling as potential targets for therapeutic intervention in FTD.
Keywords: ELAVL4; MAPT; autophagy; frontotemporal dementia; glutamatergic neurons; organoids; splicing; synaptic signaling; tauopathy.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests J.D.L. employee, Amgen. A.M.G.: Scientific Advisory Board (SAB), Denali Therapeutics (2015–2018); Genetic SAB, Pfizer (2019); SAB, Genentech; consultant, GSK, AbbVie, Biogen, and Eisai. S.J.H.: SAB, Rodin Therapeutics, Frequency Therapeutics, Psy Therapeutics, Vesigen Therapeutics, and Souvien Therapeutics; inventor, patent 6,475,723. S.T.: president, StemCultures; cofounder, LUXA Biotech; SAB, Sana Biotechnology, Blue Rock Therapeutics, and Vita Therapeutics; inventor, patent 16/331,063. J.K.I.: cofounder, AcuraStem and Modulo Bio; SAB, Spinogenix. Named companies were not involved in this project.
Figures
Comment in
-
Modeling neurodegeneration with mutant-tau organoids.Cell. 2021 Aug 19;184(17):4377-4379. doi: 10.1016/j.cell.2021.07.031. Cell. 2021. PMID: 34416145
References
Publication types
MeSH terms
Substances
Grants and funding
- P30 AG066444/AG/NIA NIH HHS/United States
- R01 NS110890/NS/NINDS NIH HHS/United States
- F31 NS117075/NS/NINDS NIH HHS/United States
- R35 NS097277/NS/NINDS NIH HHS/United States
- R01 NS095252/NS/NINDS NIH HHS/United States
- U01 AG045390/AG/NIA NIH HHS/United States
- U54 NS092089/NS/NINDS NIH HHS/United States
- R56 NS110890/NS/NINDS NIH HHS/United States
- R01 AG054008/AG/NIA NIH HHS/United States
- R01 AG056293/AG/NIA NIH HHS/United States
- P50 AG005681/AG/NIA NIH HHS/United States
- S10 OD026880/OD/NIH HHS/United States
- R01 NS097850/NS/NINDS NIH HHS/United States
- K01 AG046374/AG/NIA NIH HHS/United States
- RF1 NS110890/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
