

TNPO2 variants associate with human developmental delays, neurologic deficits, and dysmorphic features and alter TNPO2 activity in Drosophila
- PMID: 34314705
- PMCID: PMC8456166
- DOI: 10.1016/j.ajhg.2021.06.019
TNPO2 variants associate with human developmental delays, neurologic deficits, and dysmorphic features and alter TNPO2 activity in Drosophila
Abstract
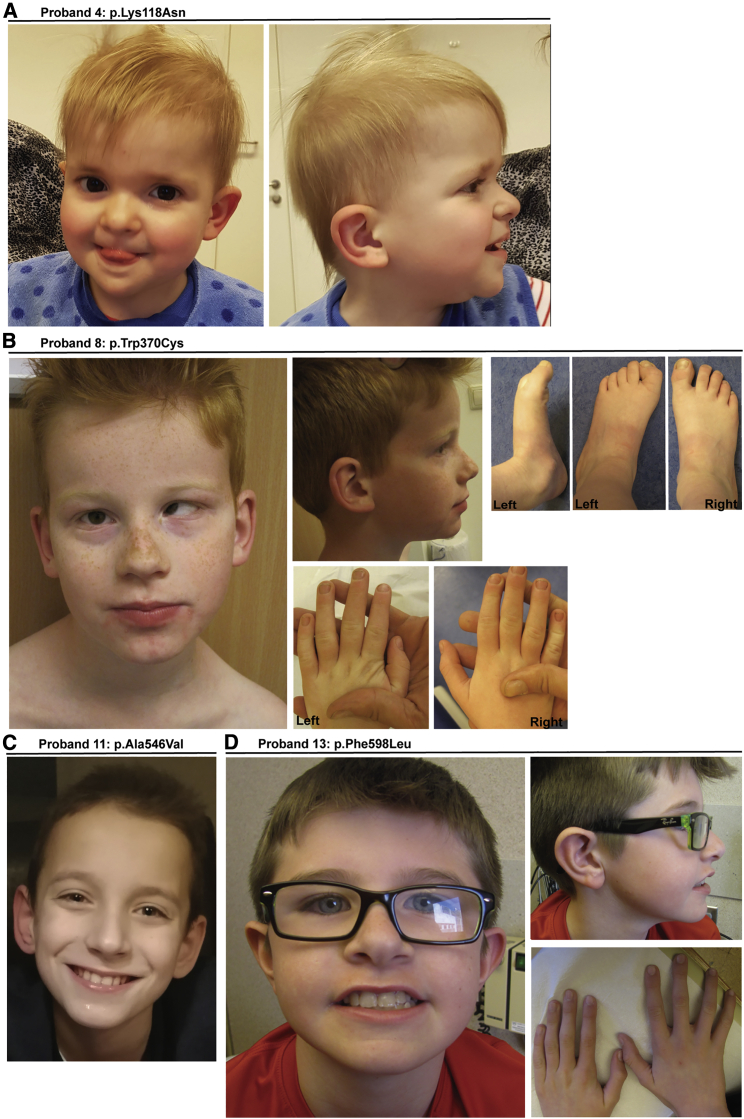
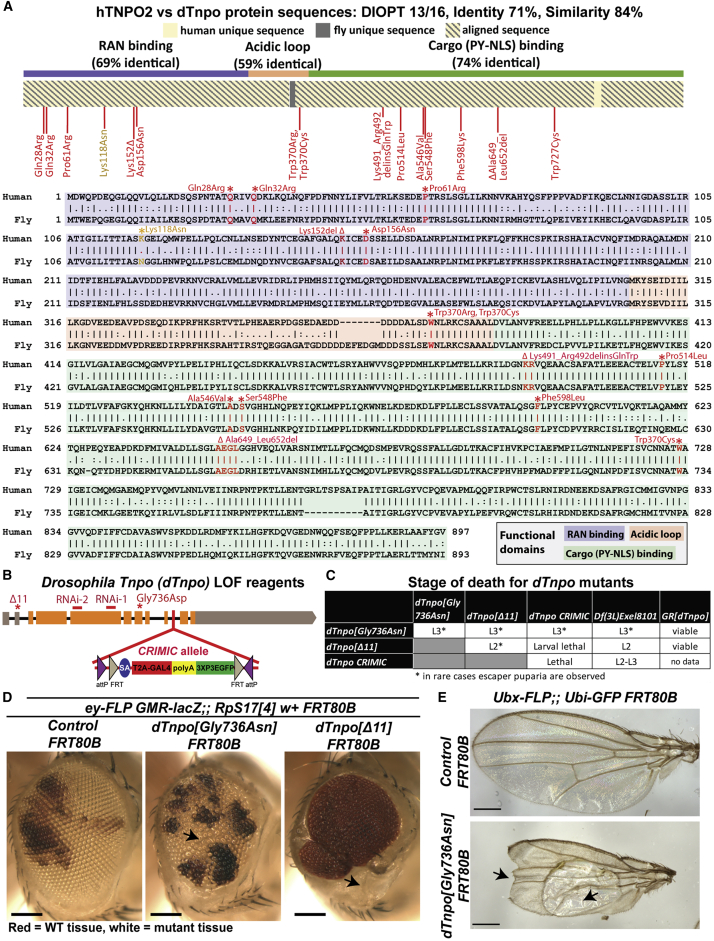
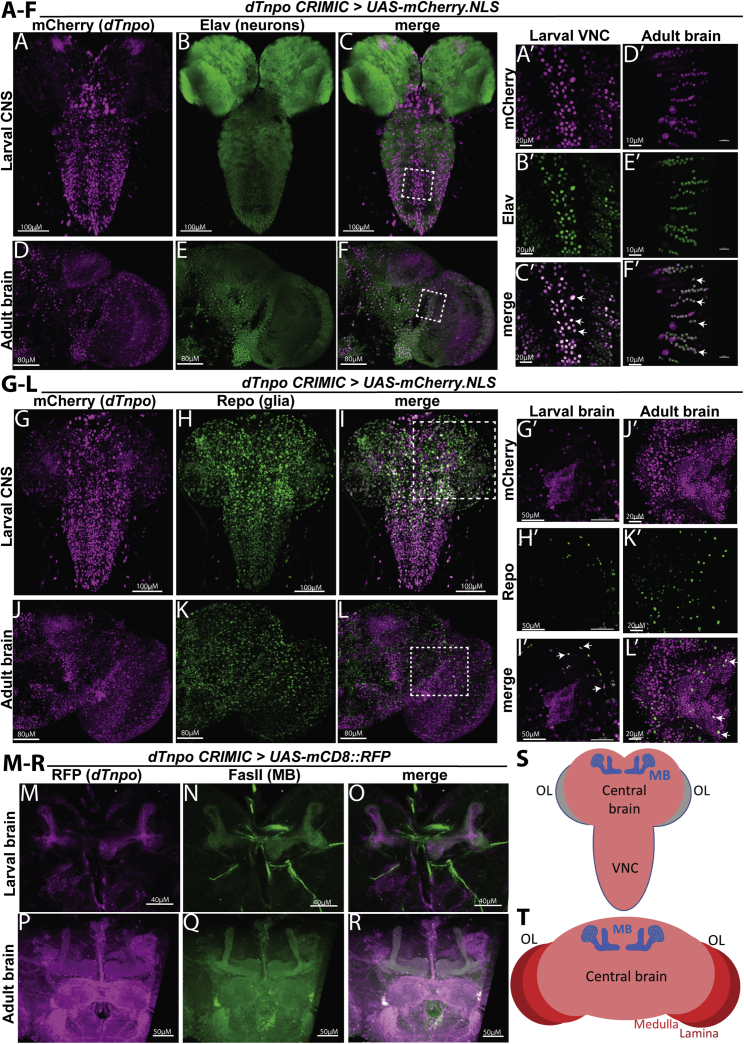
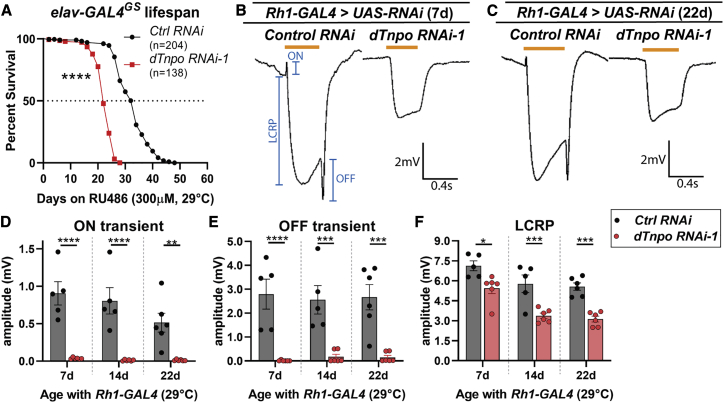
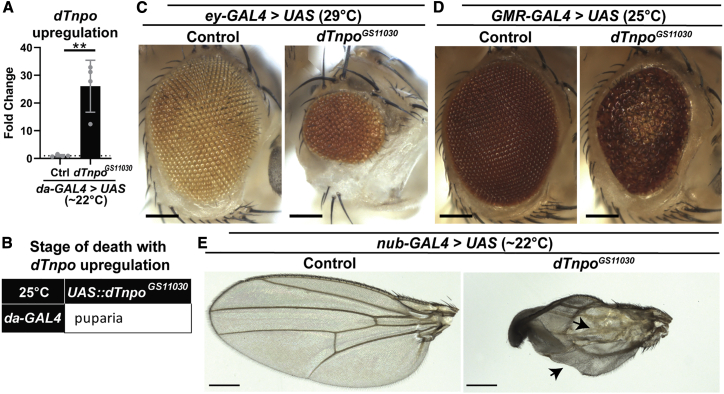
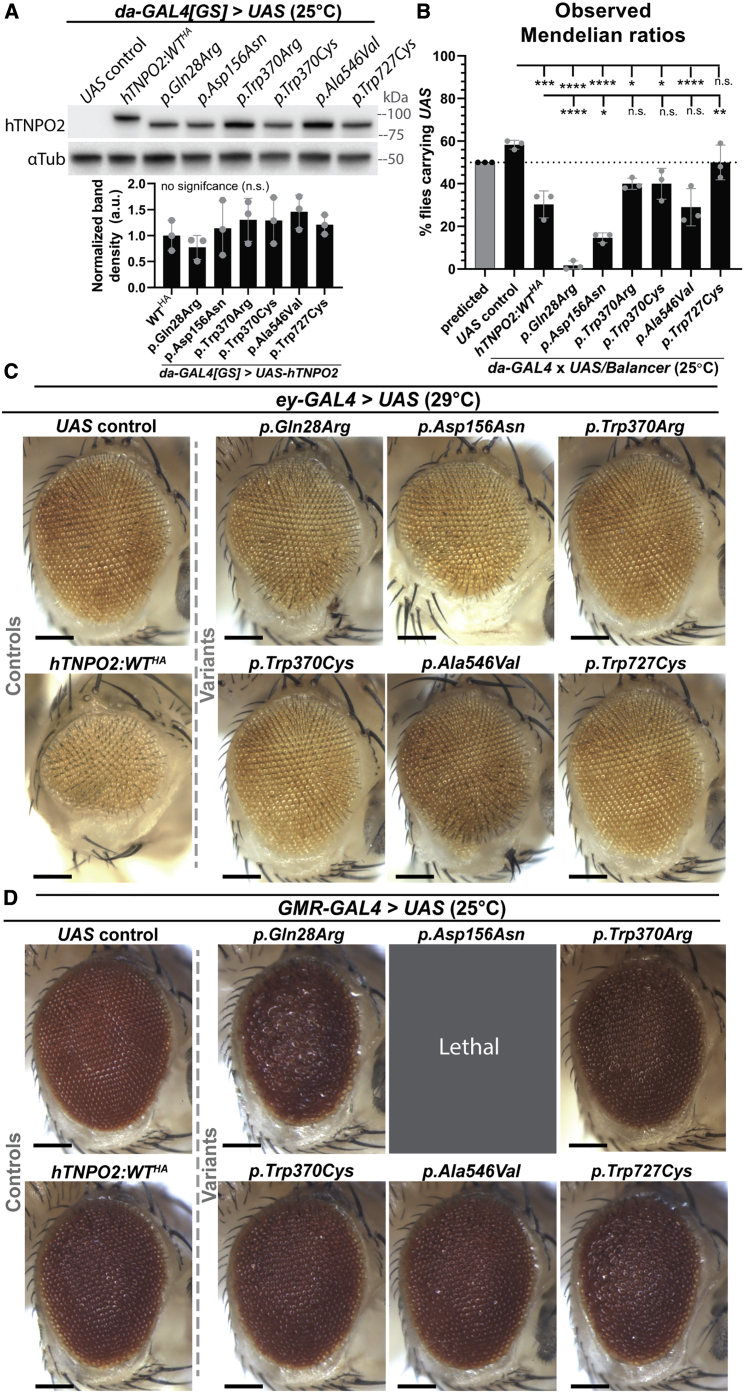
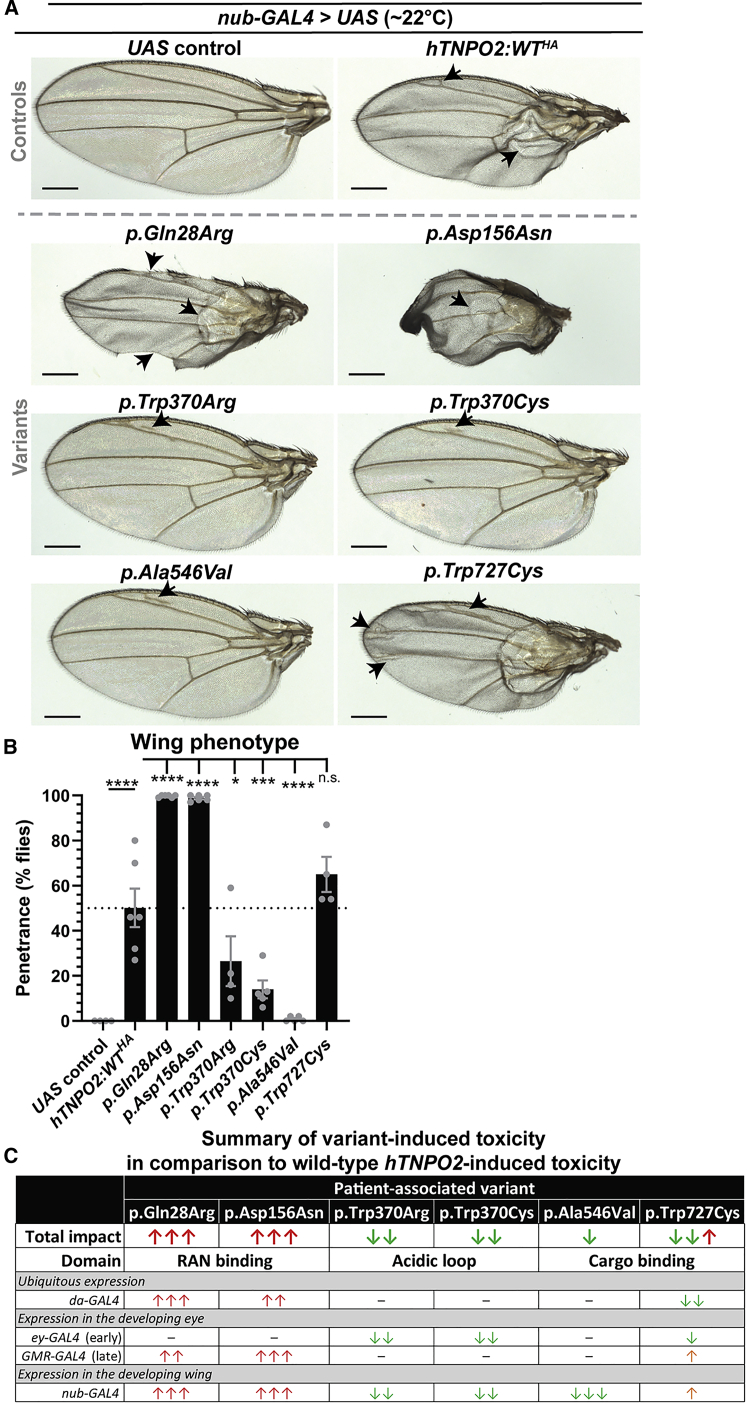
Transportin-2 (TNPO2) mediates multiple pathways including non-classical nucleocytoplasmic shuttling of >60 cargoes, such as developmental and neuronal proteins. We identified 15 individuals carrying de novo coding variants in TNPO2 who presented with global developmental delay (GDD), dysmorphic features, ophthalmologic abnormalities, and neurological features. To assess the nature of these variants, functional studies were performed in Drosophila. We found that fly dTnpo (orthologous to TNPO2) is expressed in a subset of neurons. dTnpo is critical for neuronal maintenance and function as downregulating dTnpo in mature neurons using RNAi disrupts neuronal activity and survival. Altering the activity and expression of dTnpo using mutant alleles or RNAi causes developmental defects, including eye and wing deformities and lethality. These effects are dosage dependent as more severe phenotypes are associated with stronger dTnpo loss. Interestingly, similar phenotypes are observed with dTnpo upregulation and ectopic expression of TNPO2, showing that loss and gain of Transportin activity causes developmental defects. Further, proband-associated variants can cause more or less severe developmental abnormalities compared to wild-type TNPO2 when ectopically expressed. The impact of the variants tested seems to correlate with their position within the protein. Specifically, those that fall within the RAN binding domain cause more severe toxicity and those in the acidic loop are less toxic. Variants within the cargo binding domain show tissue-dependent effects. In summary, dTnpo is an essential gene in flies during development and in neurons. Further, proband-associated de novo variants within TNPO2 disrupt the function of the encoded protein. Hence, TNPO2 variants are causative for neurodevelopmental abnormalities.
Keywords: Drosophila; Importin-3; Karyopherin-β2b; TNPO1; TNPO2; Transportin; global developmental delays; intellectual disability; nucleocytoplasmic shuttling; rare disease.
Copyright © 2021 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The Department of Molecular and Human Genetics at Baylor College of Medicine receives revenue from clinical genetic testing completed at Baylor Genetics Laboratories. Y.S. and A.B. are employees of GeneDx, Inc.
Figures
References
-
- Wangler M.F., Yamamoto S., Chao H.-T., Posey J.E., Westerfield M., Postlethwait J., Hieter P., Boycott K.M., Campeau P.M., Bellen H.J., Members of the Undiagnosed Diseases Network (UDN) Model Organisms Facilitate Rare Disease Diagnosis and Therapeutic Research. Genetics. 2017;207:9–27. - PMC - PubMed
-
- Twyffels L., Gueydan C., Kruys V. Transportin-1 and Transportin-2: protein nuclear import and beyond. FEBS Lett. 2014;588:1857–1868. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
