

SARS-CoV-2 infection triggers widespread host mRNA decay leading to an mRNA export block
- PMID: 34315815
- PMCID: PMC8522697
- DOI: 10.1261/rna.078923.121
SARS-CoV-2 infection triggers widespread host mRNA decay leading to an mRNA export block
Abstract
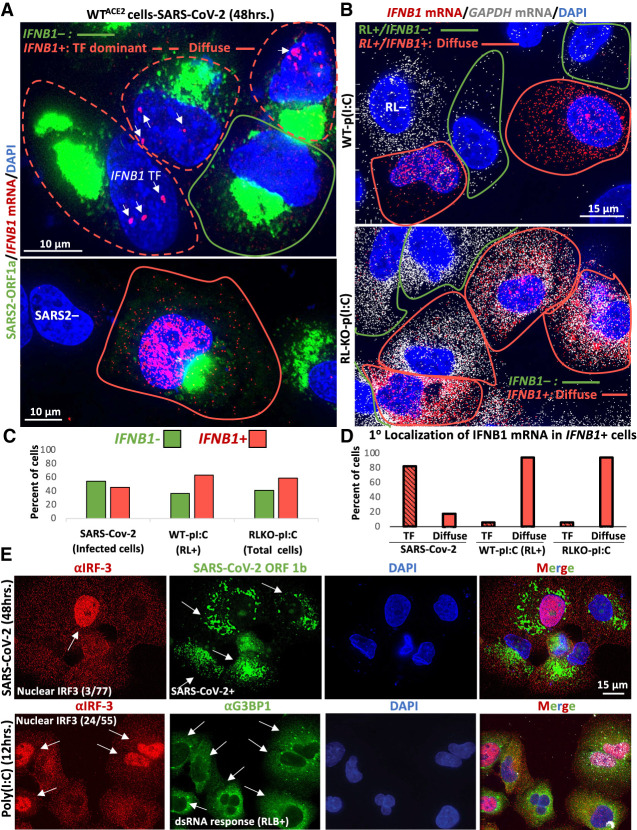
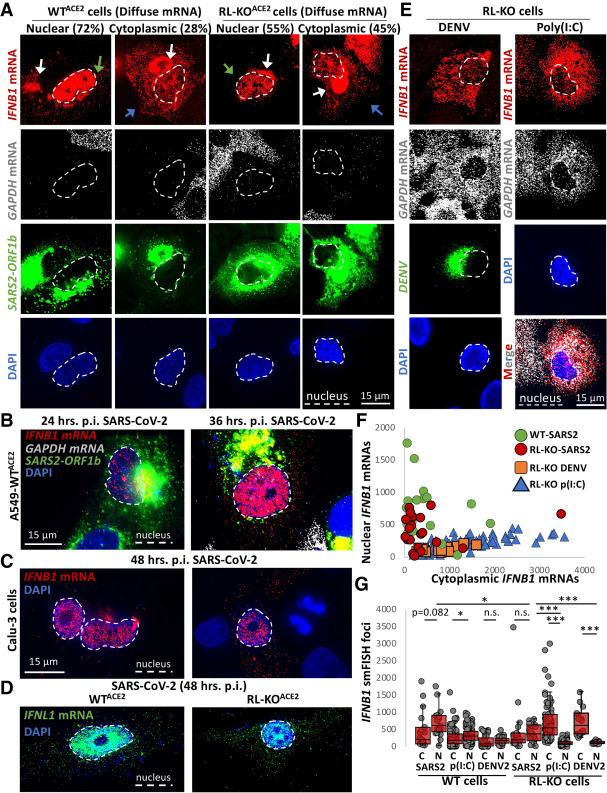
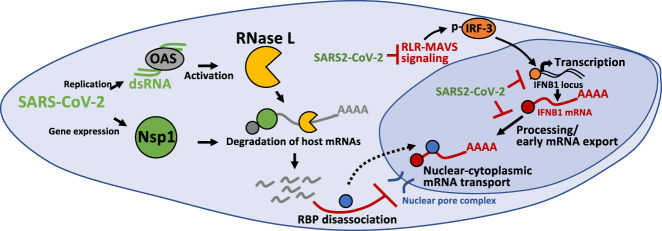
The transcriptional induction of interferon (IFN) genes is a key feature of the mammalian antiviral response that limits viral replication and dissemination. A hallmark of severe COVID-19 disease caused by SARS-CoV-2 is the low presence of IFN proteins in patient serum despite elevated levels of IFN-encoding mRNAs, indicative of post-transcriptional inhibition of IFN protein production. Here, we performed single-molecule RNA visualization to examine the expression and localization of host mRNAs during SARS-CoV-2 infection. Our data show that the biogenesis of type I and type III IFN mRNAs is inhibited at multiple steps during SARS-CoV-2 infection. First, translocation of the interferon regulatory factor 3 (IRF3) transcription factor to the nucleus is limited in response to SARS-CoV-2, indicating that SARS-CoV-2 inhibits RLR-MAVS signaling and thus weakens transcriptional induction of IFN genes. Second, we observed that IFN mRNAs primarily localize to the site of transcription in most SARS-CoV-2 infected cells, suggesting that SARS-CoV-2 either inhibits the release of IFN mRNAs from their sites of transcription and/or triggers decay of IFN mRNAs in the nucleus upon exiting the site of transcription. Lastly, nuclear-cytoplasmic transport of IFN mRNAs is inhibited during SARS-CoV-2 infection, which we propose is a consequence of widespread degradation of host cytoplasmic basal mRNAs in the early stages of SARS-CoV-2 replication by the SARS-CoV-2 Nsp1 protein, as well as the host antiviral endoribonuclease, RNase L. Importantly, IFN mRNAs can escape SARS-CoV-2-mediated degradation if they reach the cytoplasm, making rescue of mRNA export a viable means for promoting the immune response to SARS-CoV-2.
Keywords: IRF3; RNase L; SARS-CoV-2; innate immunity; interferon; mRNA decay; mRNA export.
© 2021 Burke et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
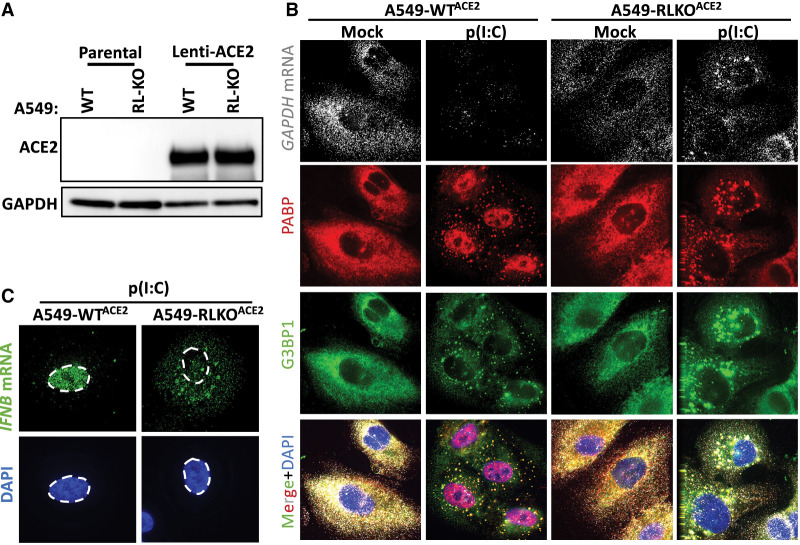
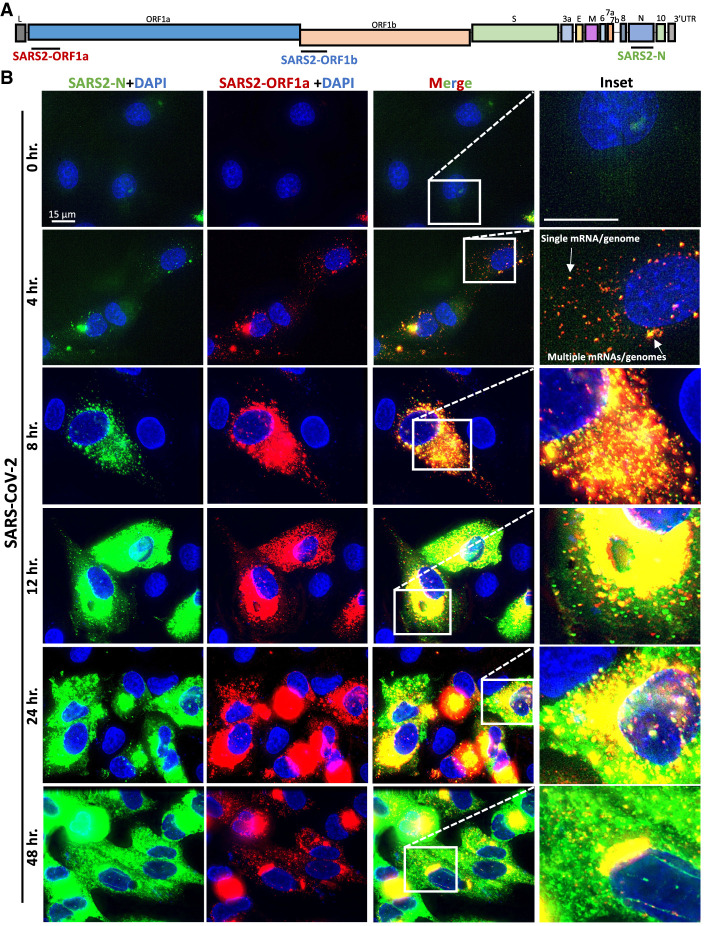
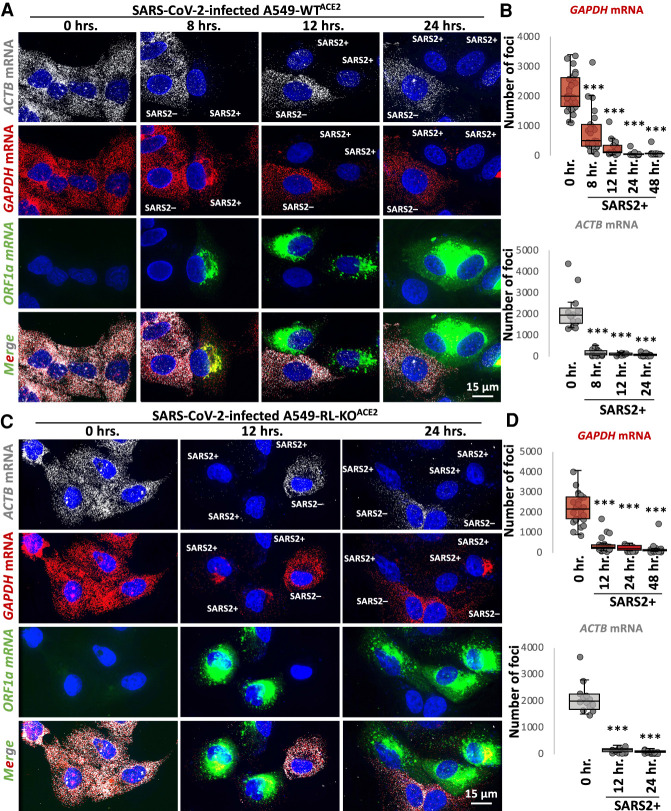
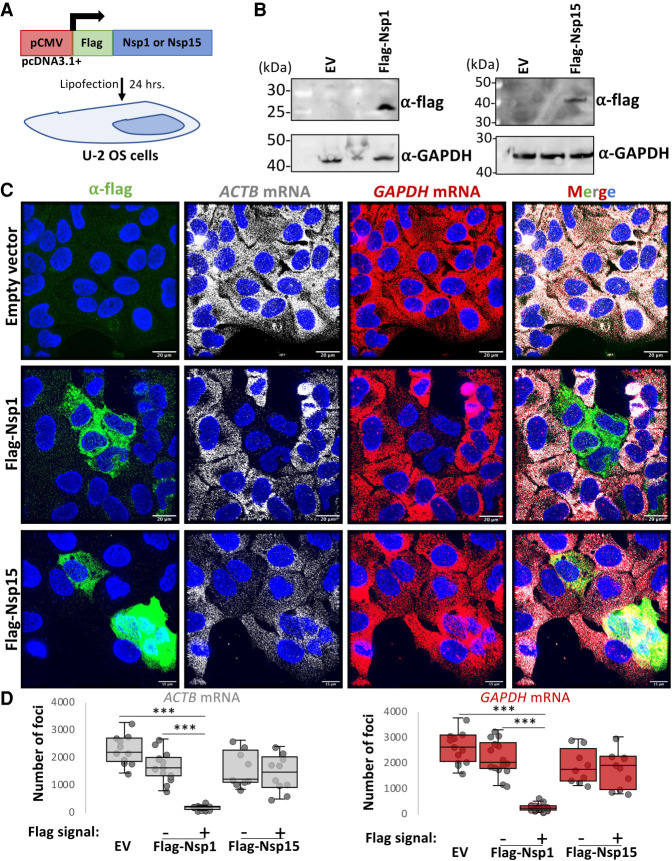
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous