

Kinetic Multi-omic Analysis of Responses to SARS-CoV-2 Infection in a Model of Severe COVID-19
- PMID: 34319784
- PMCID: PMC8475517
- DOI: 10.1128/JVI.01010-21
Kinetic Multi-omic Analysis of Responses to SARS-CoV-2 Infection in a Model of Severe COVID-19
Abstract
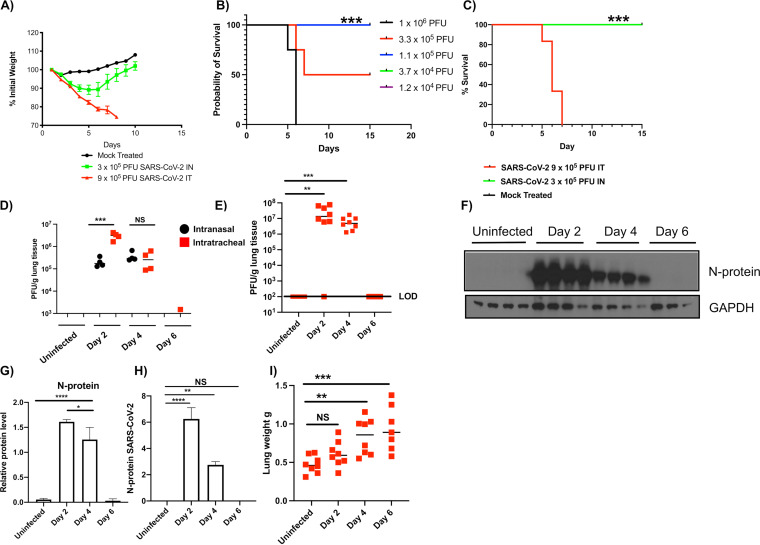
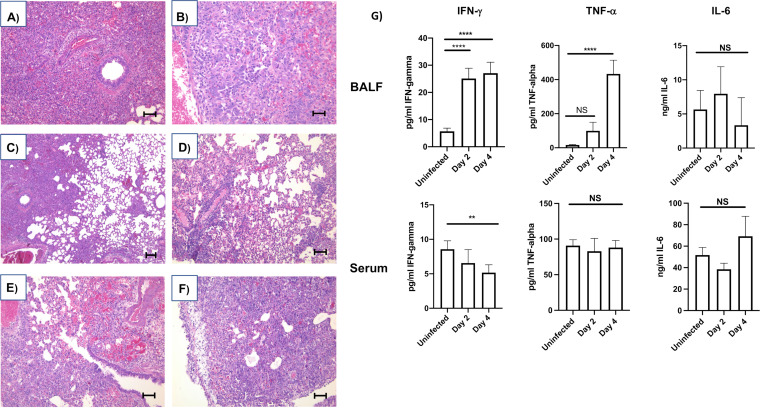
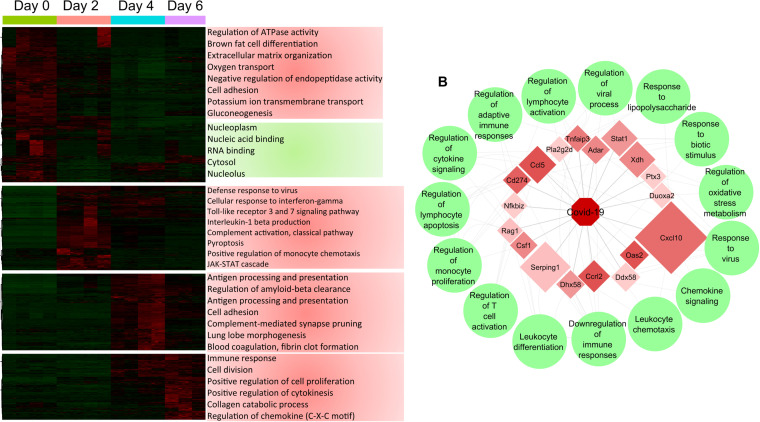
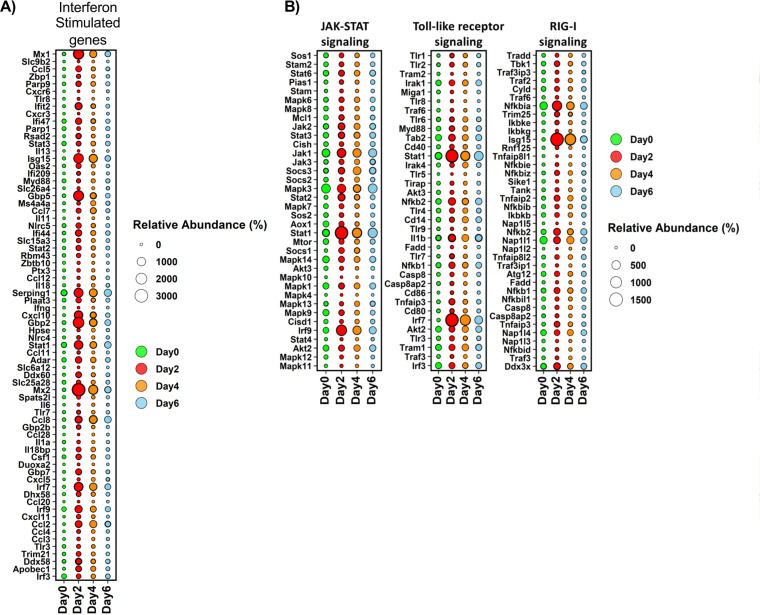
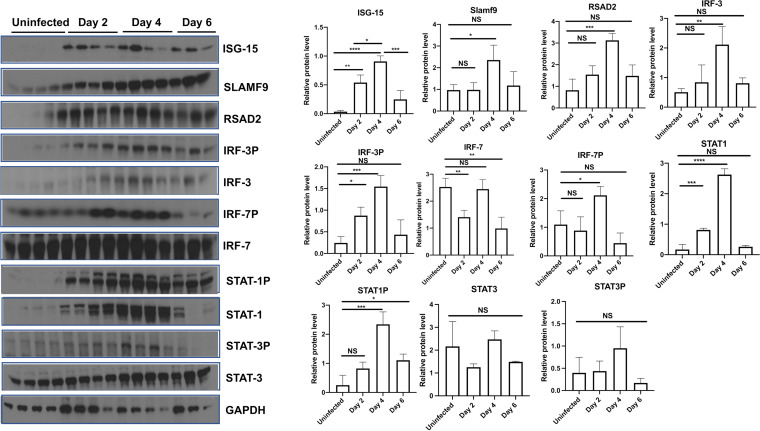
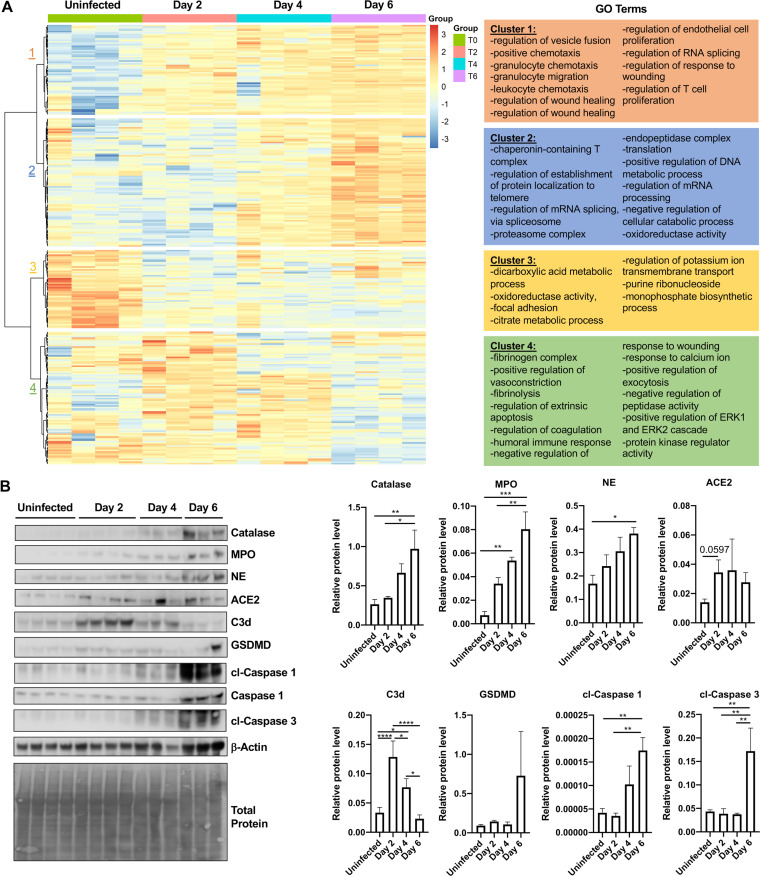
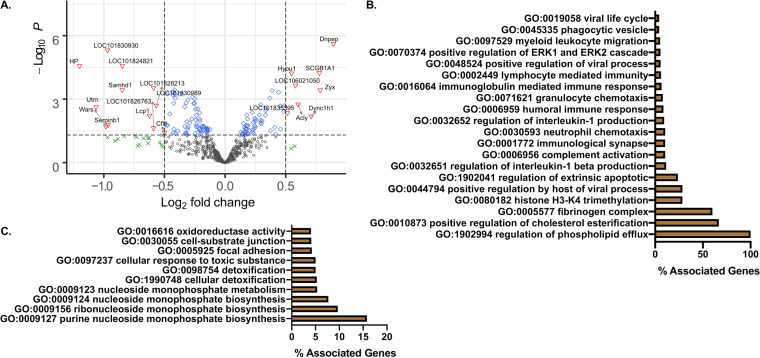
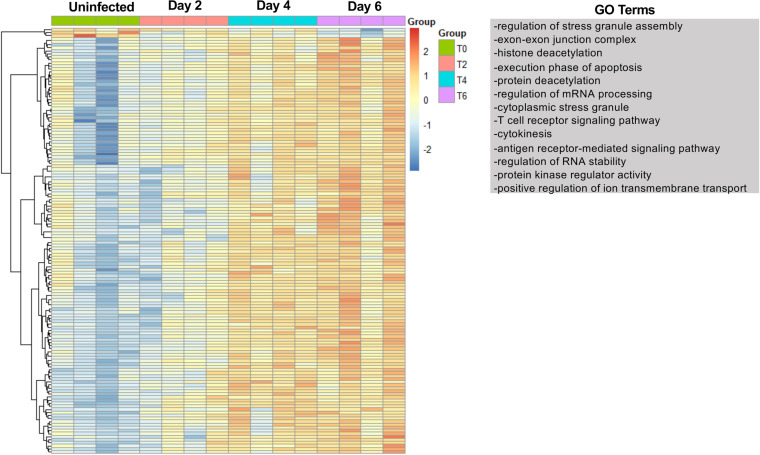
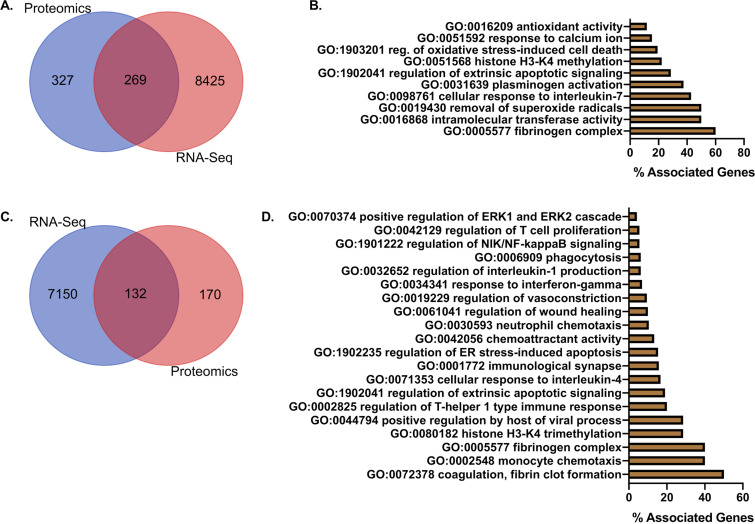
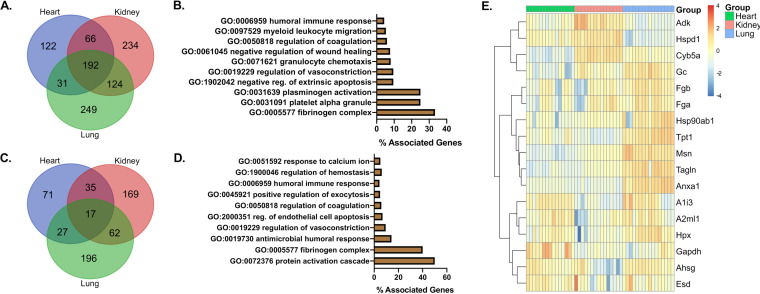
The host response to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is poorly understood due to a lack of an animal model that recapitulates severe human disease. Here, we report a Syrian hamster model that develops progressive lethal pulmonary disease that closely mimics severe coronavirus disease 2019 (COVID-19). We evaluated host responses using a multi-omic, multiorgan approach to define proteome, phosphoproteome, and transcriptome changes. These data revealed both type I and type II interferon-stimulated gene and protein expression along with a progressive increase in chemokines, monocytes, and neutrophil-associated molecules throughout the course of infection that peaked in the later time points correlating with a rapidly developing diffuse alveolar destruction and pneumonia that persisted in the absence of active viral infection. Extrapulmonary proteome and phosphoproteome remodeling was detected in the heart and kidneys following viral infection. Together, our results provide a kinetic overview of multiorgan host responses to severe SARS-CoV-2 infection in vivo. IMPORTANCE The current pandemic caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection has created an urgent need to understand the pathogenesis of this infection. These efforts have been impaired by the lack of animal models that recapitulate severe coronavirus disease 2019 (COVID-19). Here, we report a hamster model that develops severe COVID-19-like disease following infection with human isolates of SARS-CoV-2. To better understand pathogenesis, we evaluated changes in gene transcription and protein expression over the course of infection to provide an integrated multiorgan kinetic analysis of the host response to infection. These data reveal a dynamic innate immune response to infection and corresponding immune pathologies consistent with severe human disease. Altogether, this model will be useful for understanding the pathogenesis of severe COVID-19 and for testing interventions.
Keywords: COVID-19; RNA-seq; SARS-CoV-2; hamster; phosphoproteomics; proteomics.
Figures
Similar articles
-
CCR2 Signaling Restricts SARS-CoV-2 Infection.mBio. 2021 Dec 21;12(6):e0274921. doi: 10.1128/mBio.02749-21. Epub 2021 Nov 9. mBio. 2021. PMID: 34749524 Free PMC article.
-
Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures.J Virol. 2020 Sep 15;94(19):e00985-20. doi: 10.1128/JVI.00985-20. Print 2020 Sep 15. J Virol. 2020. PMID: 32699094 Free PMC article.
-
Exploring the biological mechanisms of severe COVID-19 in the elderly: Insights from an aged mouse model.Virulence. 2025 Dec;16(1):2487671. doi: 10.1080/21505594.2025.2487671. Epub 2025 Apr 14. Virulence. 2025. PMID: 40228062 Free PMC article.
-
Measures implemented in the school setting to contain the COVID-19 pandemic.Cochrane Database Syst Rev. 2022 Jan 17;1(1):CD015029. doi: 10.1002/14651858.CD015029. Cochrane Database Syst Rev. 2022. Update in: Cochrane Database Syst Rev. 2024 May 2;5:CD015029. doi: 10.1002/14651858.CD015029.pub2. PMID: 35037252 Free PMC article. Updated.
-
Antibody tests for identification of current and past infection with SARS-CoV-2.Cochrane Database Syst Rev. 2022 Nov 17;11(11):CD013652. doi: 10.1002/14651858.CD013652.pub2. Cochrane Database Syst Rev. 2022. PMID: 36394900 Free PMC article.
Cited by
-
Multiomic Investigations into Lung Health and Disease.Microorganisms. 2023 Aug 19;11(8):2116. doi: 10.3390/microorganisms11082116. Microorganisms. 2023. PMID: 37630676 Free PMC article. Review.
-
Sex differences in the cardiac stress response following SARS-CoV-2 infection of ferrets.Am J Physiol Heart Circ Physiol. 2023 Nov 1;325(5):H1153-H1167. doi: 10.1152/ajpheart.00101.2023. Epub 2023 Sep 22. Am J Physiol Heart Circ Physiol. 2023. PMID: 37737732 Free PMC article.
-
Platelet proteome reveals features of cell death, antiviral response and viral replication in covid-19.Cell Death Discov. 2022 Jul 16;8(1):324. doi: 10.1038/s41420-022-01122-1. Cell Death Discov. 2022. PMID: 35842415 Free PMC article.
-
Type I interferons and SARS-CoV-2: from cells to organisms.Curr Opin Immunol. 2022 Feb;74:172-182. doi: 10.1016/j.coi.2022.01.003. Epub 2022 Jan 25. Curr Opin Immunol. 2022. PMID: 35149239 Free PMC article. Review.
-
Comprehensive Analysis of Disease Pathology in Immunocompetent and Immunocompromised Hosts following Pulmonary SARS-CoV-2 Infection.Biomedicines. 2022 Jun 7;10(6):1343. doi: 10.3390/biomedicines10061343. Biomedicines. 2022. PMID: 35740365 Free PMC article.
References
-
- Chen G, Wu D, Guo W, Cao Y, Huang D, Wang H, Wang T, Zhang X, Chen H, Yu H, Zhang X, Zhang M, Wu S, Song J, Chen T, Han M, Li S, Luo X, Zhao J, Ning Q. 2020. Clinical and immunological features of severe and moderate coronavirus disease 2019. J Clin Invest 130:2620–2629. 10.1172/JCI137244. - DOI - PMC - PubMed
-
- Xu Z, Shi L, Wang Y, Zhang J, Huang L, Zhang C, Liu S, Zhao P, Liu H, Zhu L, Tai Y, Bai C, Gao T, Song J, Xia P, Dong J, Zhao J, Wang FS. 2020. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir Med 8:420–422. 10.1016/S2213-2600(20)30076-X. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UM1 TR004538/TR/NCATS NIH HHS/United States
- R21 AI148722/AI/NIAID NIH HHS/United States
- S10 OD021805/OD/NIH HHS/United States
- P30 AG013319/AG/NIA NIH HHS/United States
- P01 AG062413/AG/NIA NIH HHS/United States
- AI148722-01/HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R56 AG057785/AG/NIA NIH HHS/United States
- R01 AG070034/AG/NIA NIH HHS/United States
- P30 CA054174/CA/NCI NIH HHS/United States
- P30-AG013319/HHS | NIH | National Institute on Aging (NIA)
- I01 BX001023/BX/BLRD VA/United States
- UL1 TR002645/TR/NCATS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
