

Targeting Mycobacterium tuberculosis response to environmental cues for the development of effective antitubercular drugs
- PMID: 34319985
- PMCID: PMC8351955
- DOI: 10.1371/journal.pbio.3001355
Targeting Mycobacterium tuberculosis response to environmental cues for the development of effective antitubercular drugs
Abstract
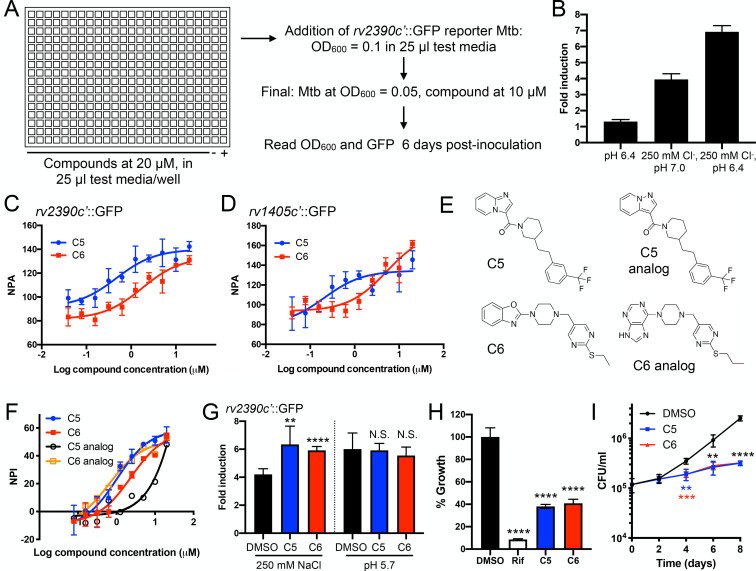
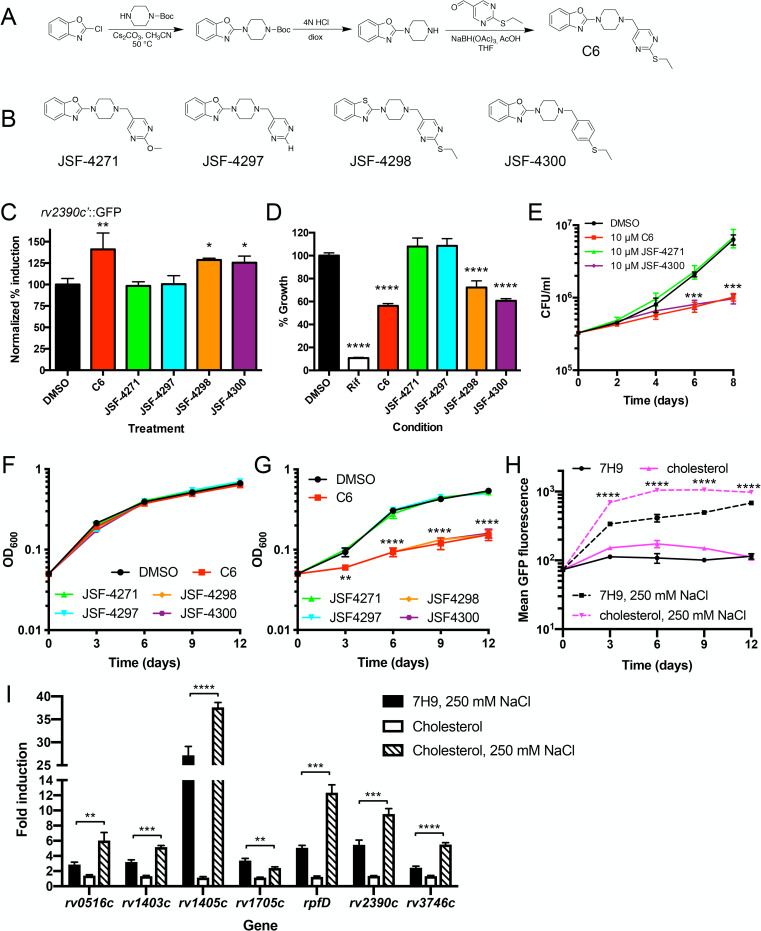
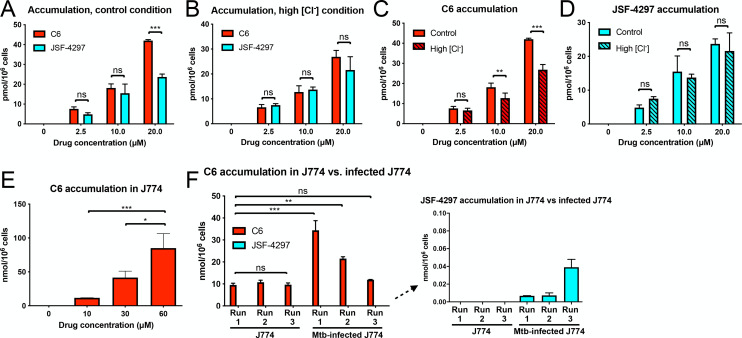
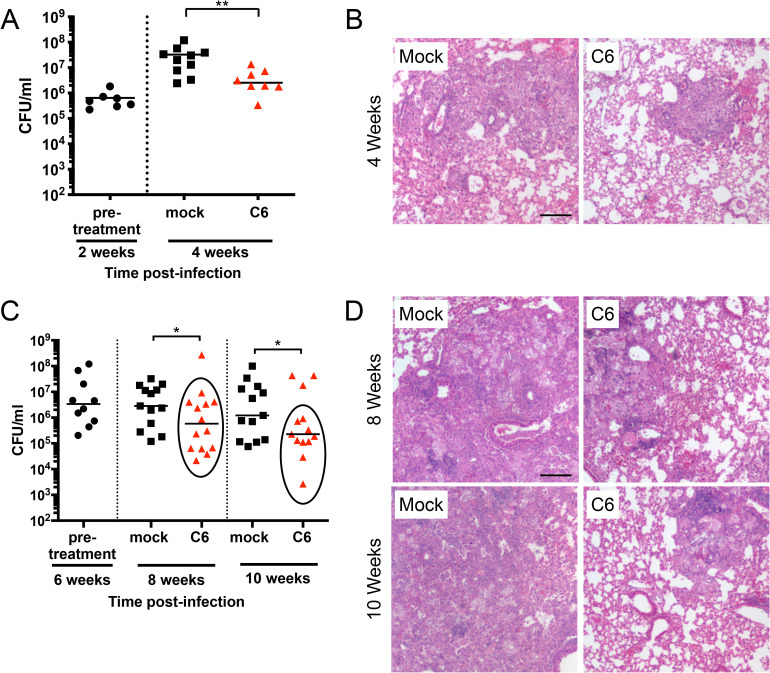
Sensing and response to environmental cues, such as pH and chloride (Cl-), is critical in enabling Mycobacterium tuberculosis (Mtb) colonization of its host. Utilizing a fluorescent reporter Mtb strain in a chemical screen, we have identified compounds that dysregulate Mtb response to high Cl- levels, with a subset of the hits also inhibiting Mtb growth in host macrophages. Structure-activity relationship studies on the hit compound "C6," or 2-(4-((2-(ethylthio)pyrimidin-5-yl)methyl)piperazin-1-yl)benzo[d]oxazole, demonstrated a correlation between compound perturbation of Mtb Cl- response and inhibition of bacterial growth in macrophages. C6 accumulated in both bacterial and host cells, and inhibited Mtb growth in cholesterol media, but not in rich media. Subsequent examination of the Cl- response of Mtb revealed an intriguing link with bacterial growth in cholesterol, with increased transcription of several Cl--responsive genes in the simultaneous presence of cholesterol and high external Cl- concentration, versus transcript levels observed during exposure to high external Cl- concentration alone. Strikingly, oral administration of C6 was able to inhibit Mtb growth in vivo in a C3HeB/FeJ murine infection model. Our work illustrates how Mtb response to environmental cues can intersect with its metabolism and be exploited in antitubercular drug discovery.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: MS, JSP, JSF, and ST are listed as inventors on a patent filing pertinent to the compounds disclosed in this manuscript as employees of Rutgers University (MS, JSP, JSF) or Tufts University (ST). All other authors have no competing interests.
Figures
Similar articles
-
Mycobacterium tuberculosis responds to chloride and pH as synergistic cues to the immune status of its host cell.PLoS Pathog. 2013;9(4):e1003282. doi: 10.1371/journal.ppat.1003282. Epub 2013 Apr 4. PLoS Pathog. 2013. PMID: 23592993 Free PMC article.
-
Novel inhibitors of cholesterol degradation in Mycobacterium tuberculosis reveal how the bacterium's metabolism is constrained by the intracellular environment.PLoS Pathog. 2015 Feb 12;11(2):e1004679. doi: 10.1371/journal.ppat.1004679. eCollection 2015 Feb. PLoS Pathog. 2015. PMID: 25675247 Free PMC article.
-
Targeting Mycobacterium tuberculosis pH-driven adaptation.Microbiology (Reading). 2024 May;170(5):001458. doi: 10.1099/mic.0.001458. Microbiology (Reading). 2024. PMID: 38717801 Free PMC article. Review.
-
Mycobacterium tuberculosis response to cholesterol is integrated with environmental pH and potassium levels via a lipid metabolism regulator.PLoS Genet. 2024 Jan 24;20(1):e1011143. doi: 10.1371/journal.pgen.1011143. eCollection 2024 Jan. PLoS Genet. 2024. PMID: 38266039 Free PMC article.
-
The impact of Mycobacterium tuberculosis on the macrophage cholesterol metabolism pathway.Front Immunol. 2024 May 30;15:1402024. doi: 10.3389/fimmu.2024.1402024. eCollection 2024. Front Immunol. 2024. PMID: 38873598 Free PMC article. Review.
Cited by
-
Rv0500A is a transcription factor that links Mycobacterium tuberculosis environmental response with division and impacts host colonization.Mol Microbiol. 2022 May;117(5):1048-1062. doi: 10.1111/mmi.14886. Epub 2022 Feb 28. Mol Microbiol. 2022. PMID: 35167150 Free PMC article.
-
Differentiating Peromyscus leucopus bone marrow-derived macrophages for characterization of responses to Borrelia burgdorferi and lipopolysaccharide.Infect Immun. 2025 Jul 8;93(7):e0058124. doi: 10.1128/iai.00581-24. Epub 2025 May 27. Infect Immun. 2025. PMID: 40422003 Free PMC article.
-
Cholesterol metabolism and intrabacterial potassium homeostasis are intrinsically related in Mycobacterium tuberculosis.PLoS Pathog. 2025 May 22;21(5):e1013207. doi: 10.1371/journal.ppat.1013207. eCollection 2025 May. PLoS Pathog. 2025. PMID: 40402977 Free PMC article.
-
Cholesterol metabolism and intrabacterial potassium homeostasis are intrinsically related in Mycobacterium tuberculosis.bioRxiv [Preprint]. 2024 Nov 11:2024.11.10.622811. doi: 10.1101/2024.11.10.622811. bioRxiv. 2024. Update in: PLoS Pathog. 2025 May 22;21(5):e1013207. doi: 10.1371/journal.ppat.1013207. PMID: 39605342 Free PMC article. Updated. Preprint.
-
PrrA modulates Mycobacterium tuberculosis response to multiple environmental cues and is critically regulated by serine/threonine protein kinases.PLoS Genet. 2022 Aug 1;18(8):e1010331. doi: 10.1371/journal.pgen.1010331. eCollection 2022 Aug. PLoS Genet. 2022. PMID: 35913986 Free PMC article.
References
-
- World Health Organization. Global tuberculosis report 2020. Available from: https://apps.who.int/iris/bitstream/handle/10665/336069/9789240013131-en... [cited 2021 Jun 4].
-
- Mattila JT, Ojo OO, Kepka-Lenhart D, Marino S, Kim JH, Eum SY, et al.. Microenvironments in tuberculous granulomas are delineated by distinct populations of macrophage subsets and expression of nitric oxide synthase and arginase isoforms. J Immunol. 2013;191(2):773–84. doi: 10.4049/jimmunol.1300113 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
