

In vivo isotope tracing reveals the versatility of glucose as a brown adipose tissue substrate
- PMID: 34320357
- PMCID: PMC8369932
- DOI: 10.1016/j.celrep.2021.109459
In vivo isotope tracing reveals the versatility of glucose as a brown adipose tissue substrate
Abstract
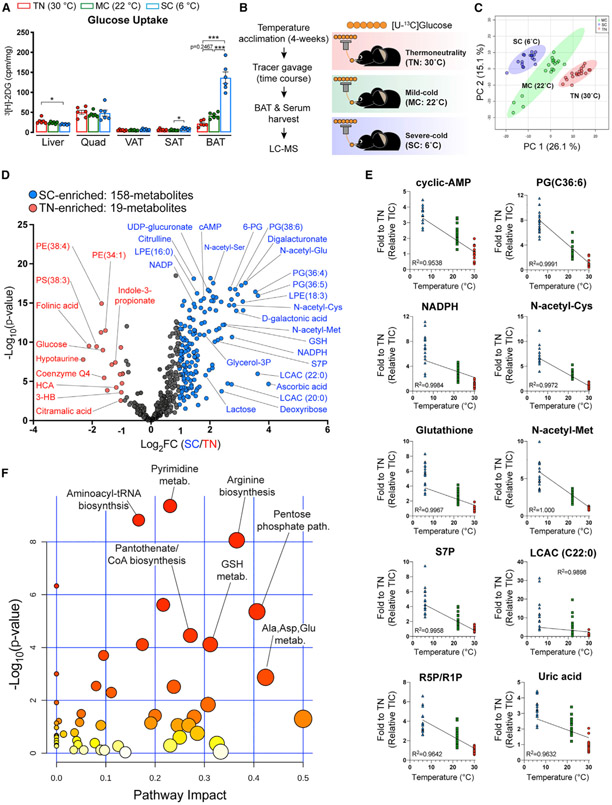
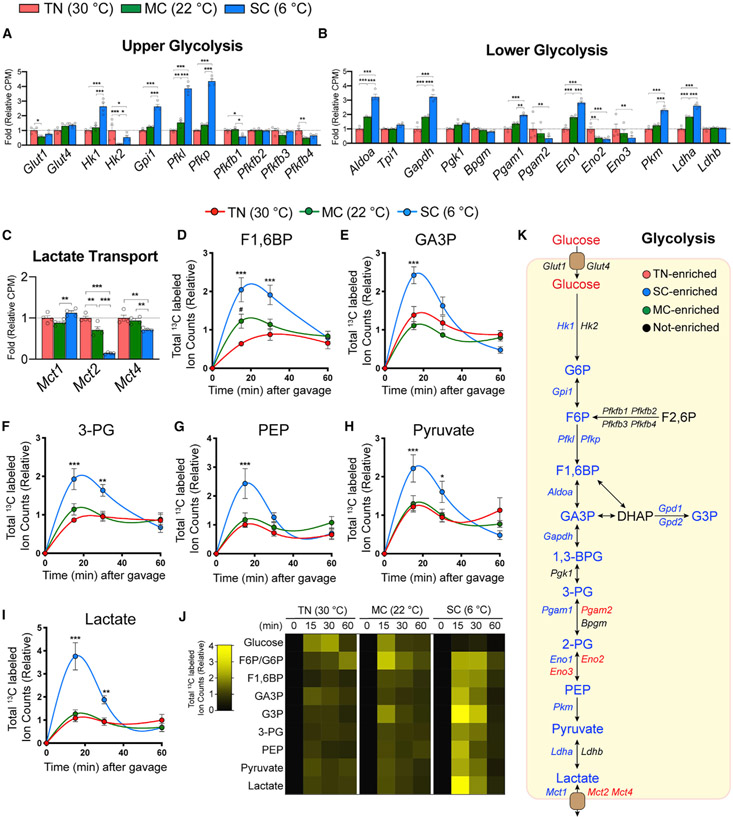
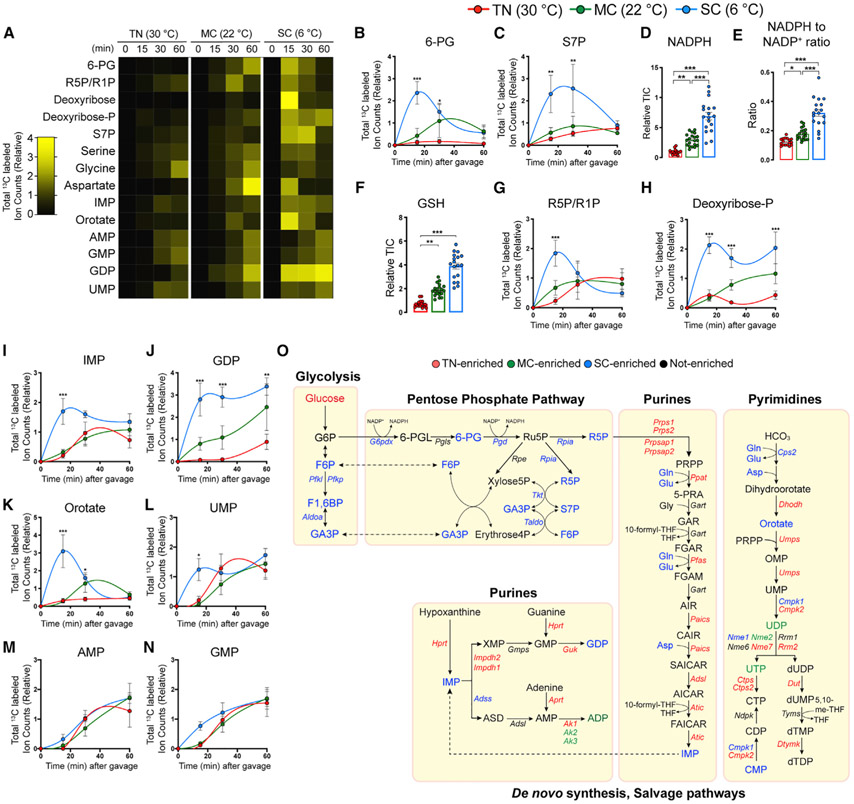
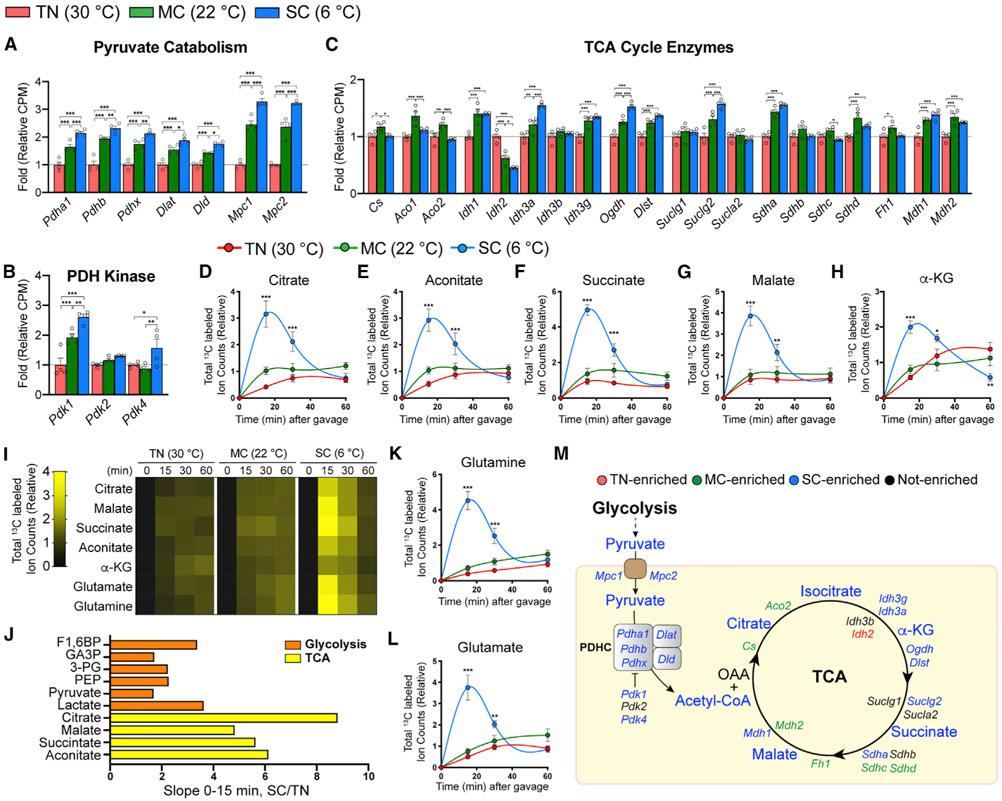
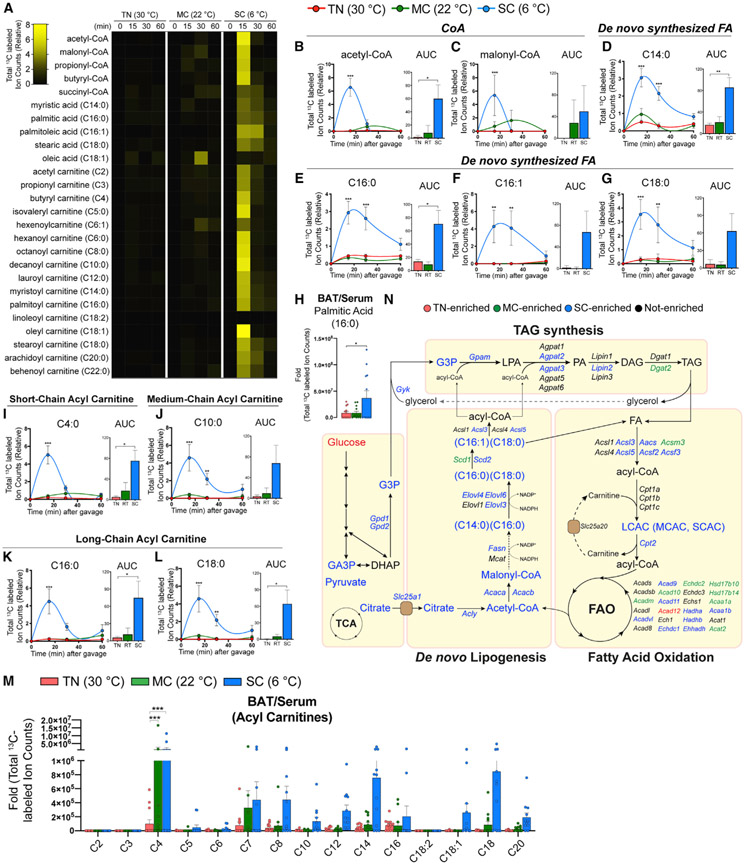
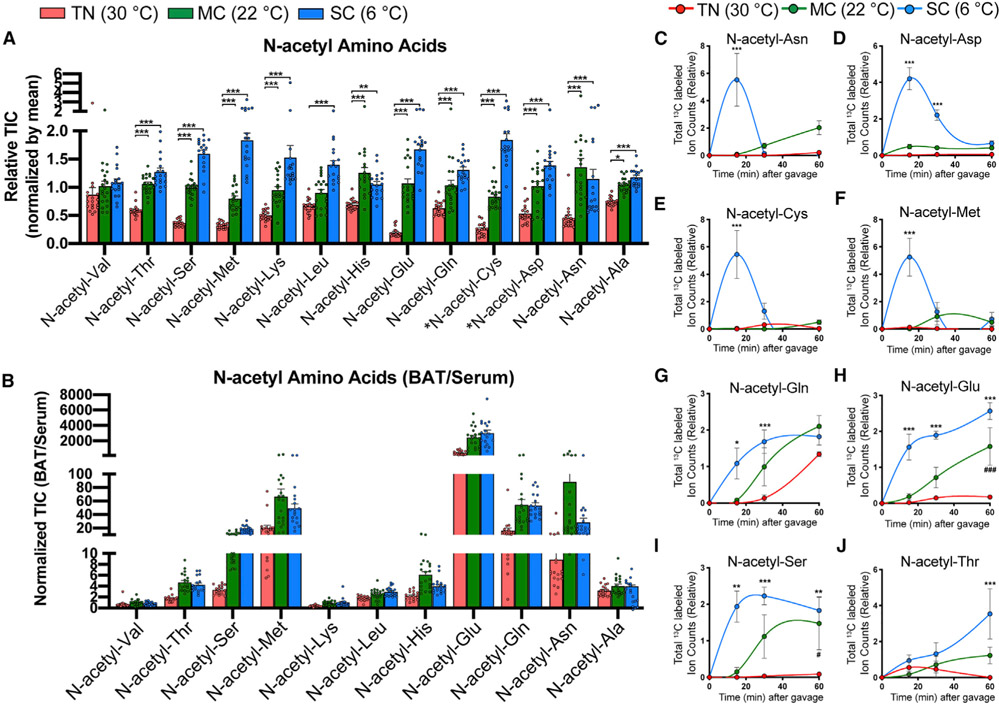
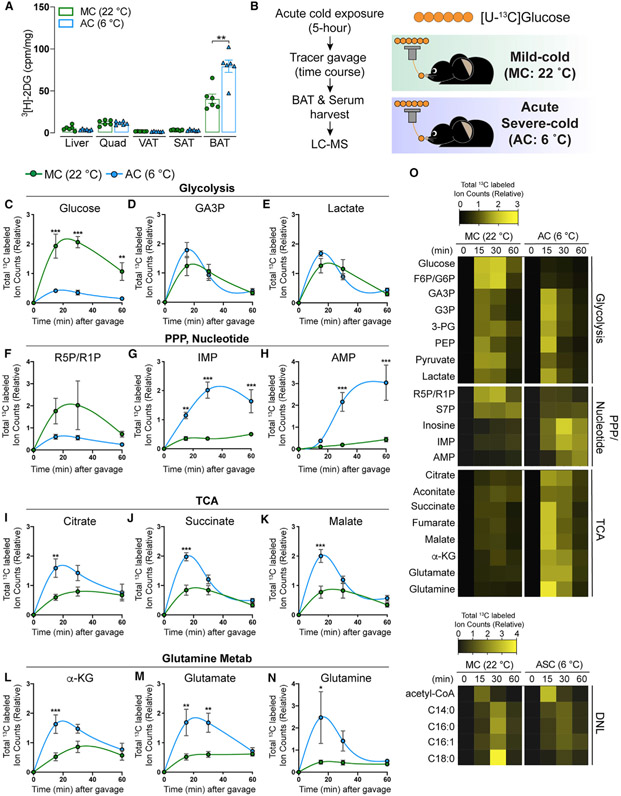
Active brown adipose tissue (BAT) consumes copious amounts of glucose, yet how glucose metabolism supports thermogenesis is unclear. By combining transcriptomics, metabolomics, and stable isotope tracing in vivo, we systematically analyze BAT glucose utilization in mice during acute and chronic cold exposure. Metabolite profiling reveals extensive temperature-dependent changes in the BAT metabolome and transcriptome upon cold adaptation, discovering unexpected metabolite markers of thermogenesis, including increased N-acetyl-amino acid production. Time-course stable isotope tracing further reveals rapid incorporation of glucose carbons into glycolysis and TCA cycle, as well as several auxiliary pathways, including NADPH, nucleotide, and phospholipid synthesis pathways. Gene expression differences inconsistently predict glucose fluxes, indicating that posttranscriptional mechanisms also govern glucose utilization. Surprisingly, BAT swiftly generates fatty acids and acyl-carnitines from glucose, suggesting that lipids are rapidly synthesized and immediately oxidized. These data reveal versatility in BAT glucose utilization, highlighting the value of an integrative-omics approach to understanding organ metabolism.
Keywords: BAT; brown adipocyte; brown adipose tissue; brown fat; glucose metabolism; lipid metabolism; metabolomics; stable isotope tracing; temperature acclimation; thermogenesis.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Allred JB, and Guy DG (1969). Determination of coenzyme A and acetyl CoA in tissue extracts. Anal. Biochem 29, 293–299. - PubMed
