

Age-related neuroinflammation and pathology in the locus coeruleus and hippocampus: beta-adrenergic antagonists exacerbate impairment of learning and memory in aged mice
- PMID: 34320462
- PMCID: PMC8419102
- DOI: 10.1016/j.neurobiolaging.2021.06.012
Age-related neuroinflammation and pathology in the locus coeruleus and hippocampus: beta-adrenergic antagonists exacerbate impairment of learning and memory in aged mice
Abstract
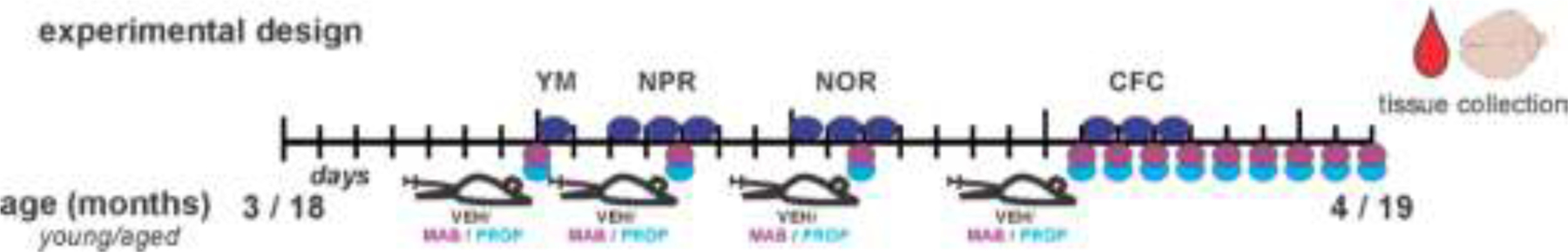
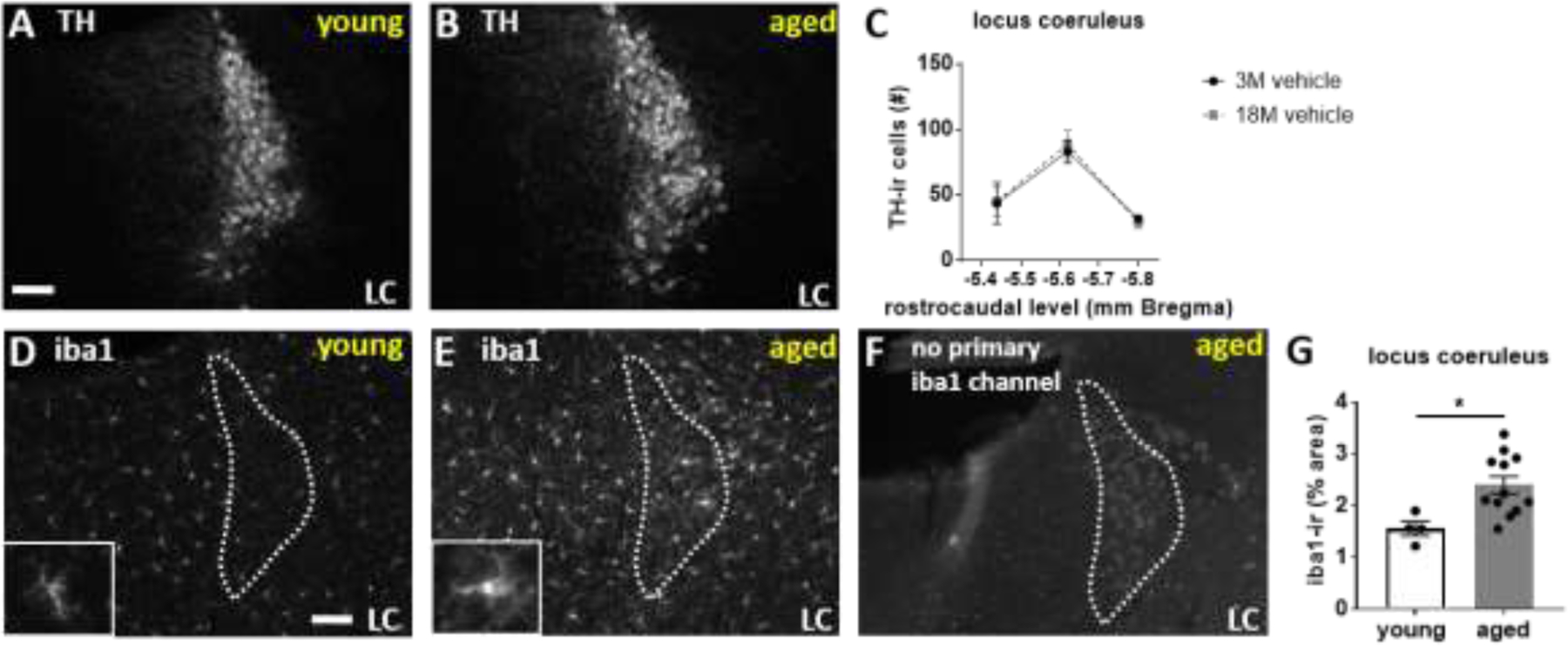
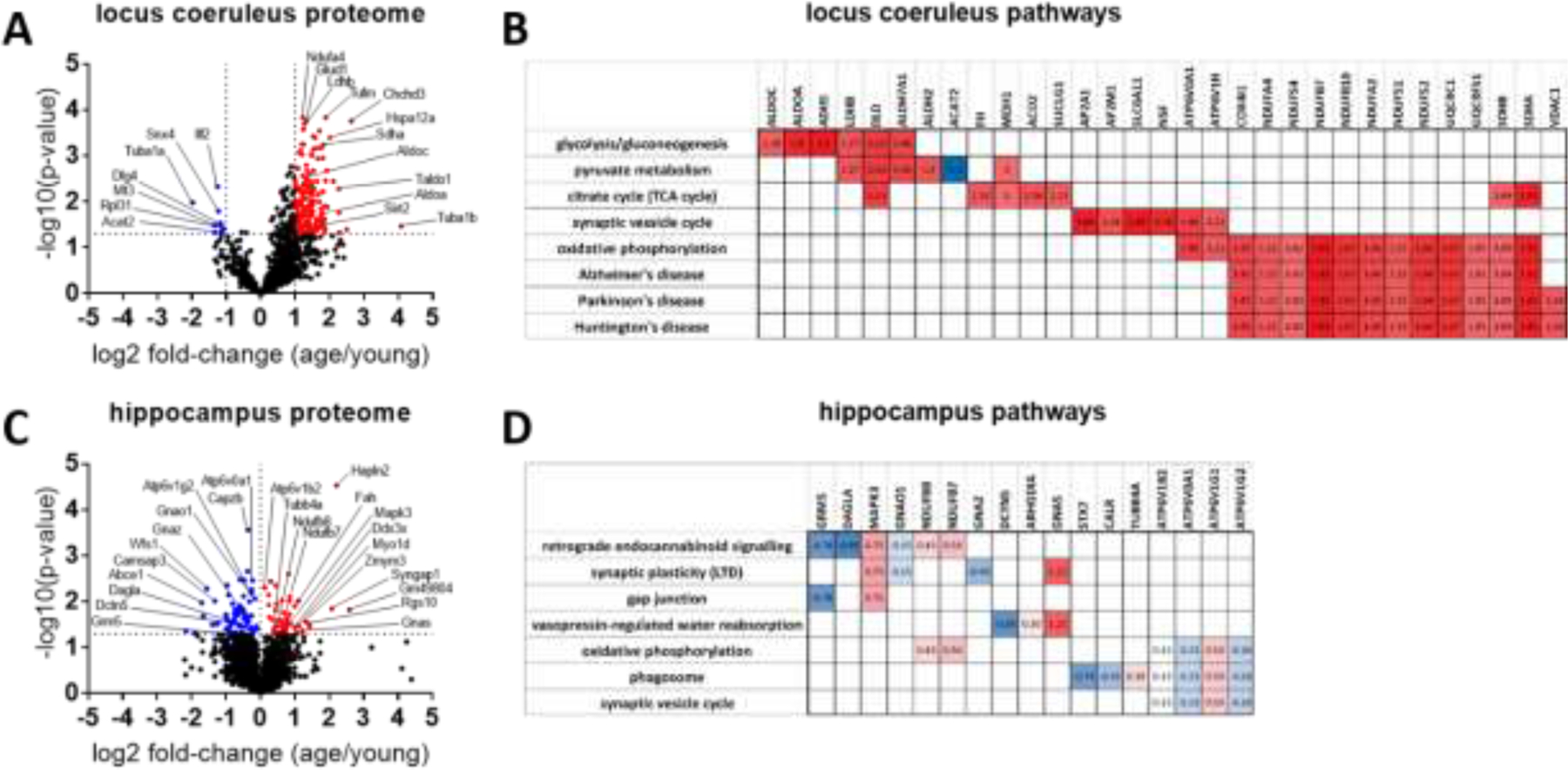
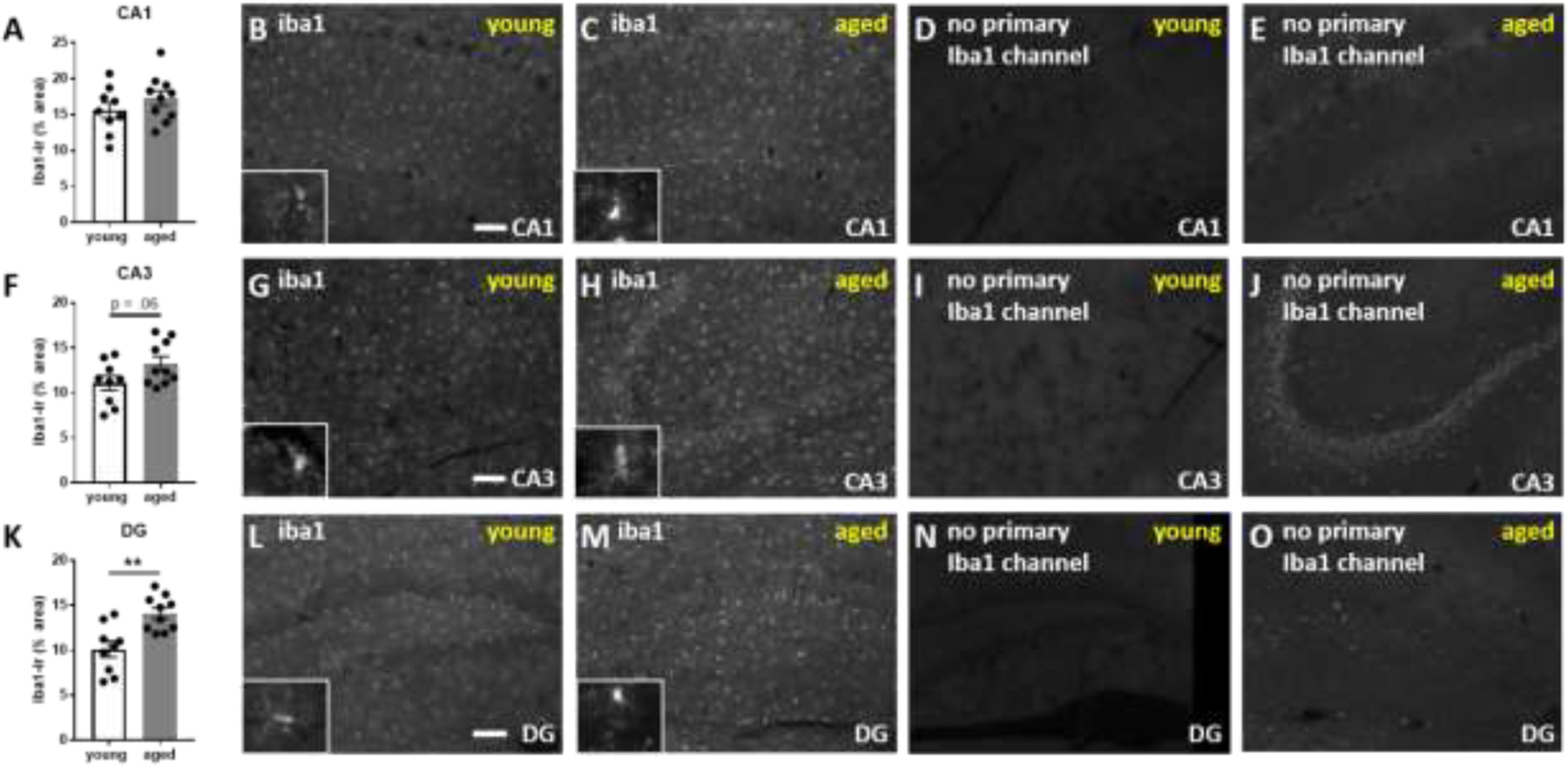
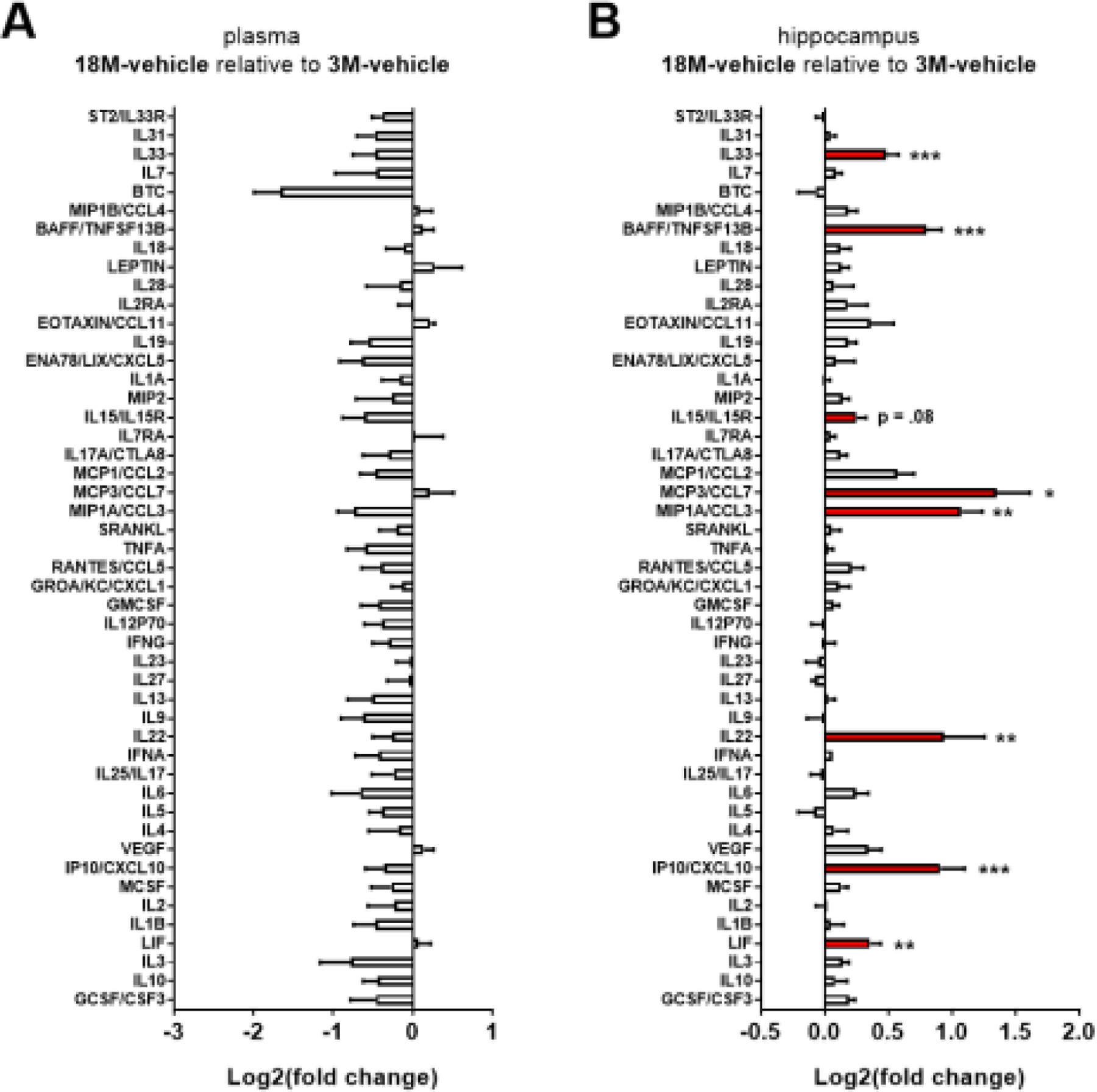
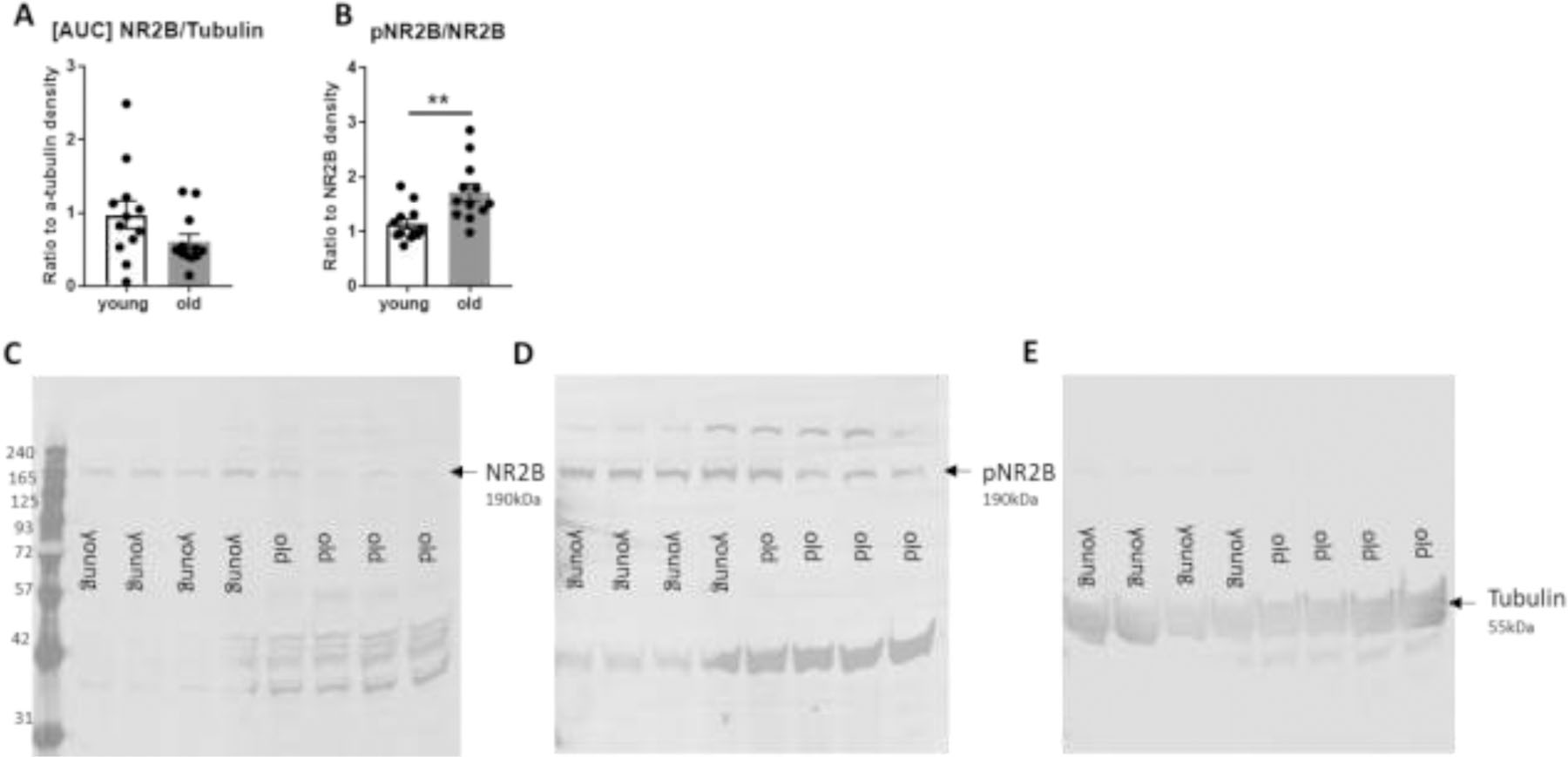
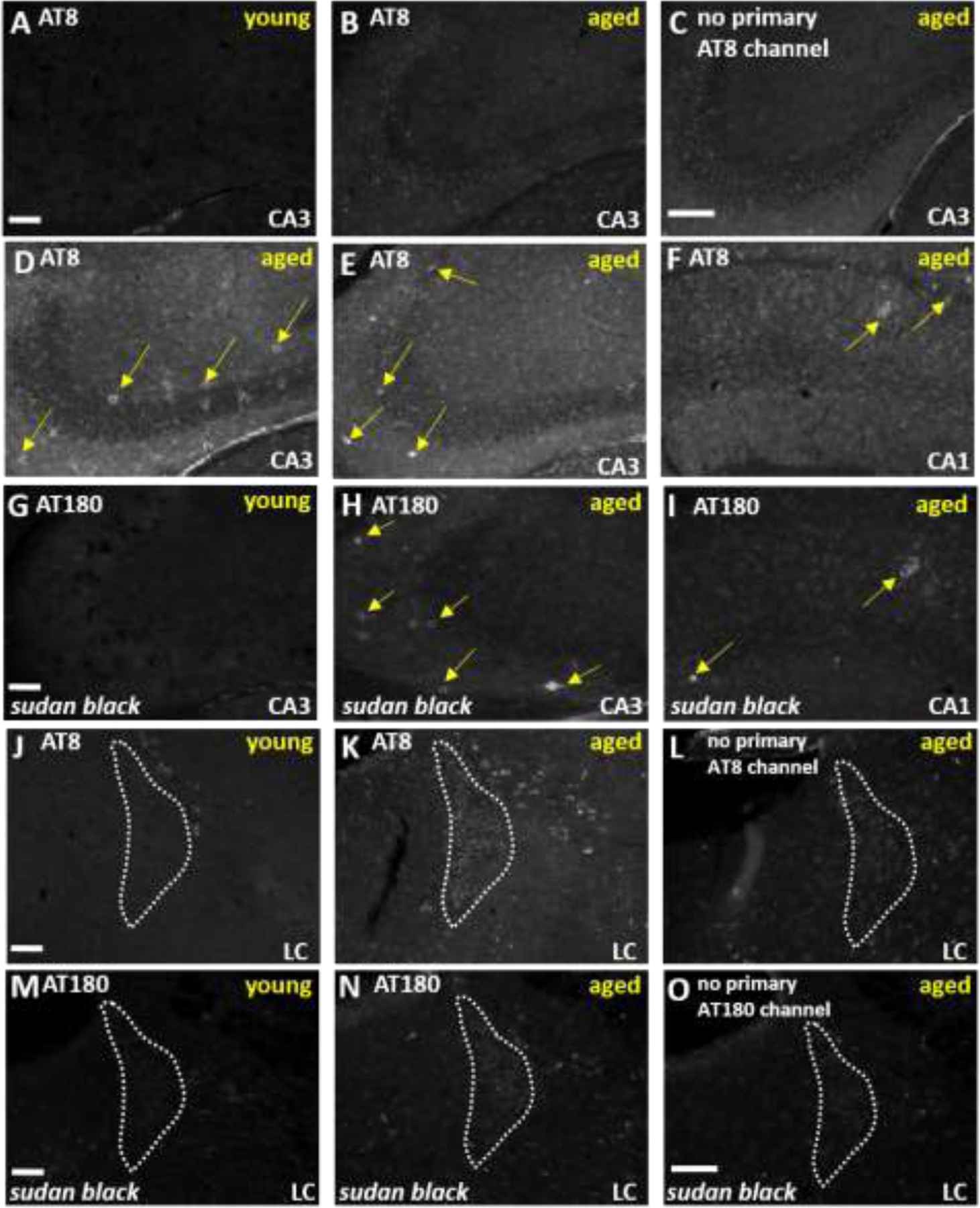
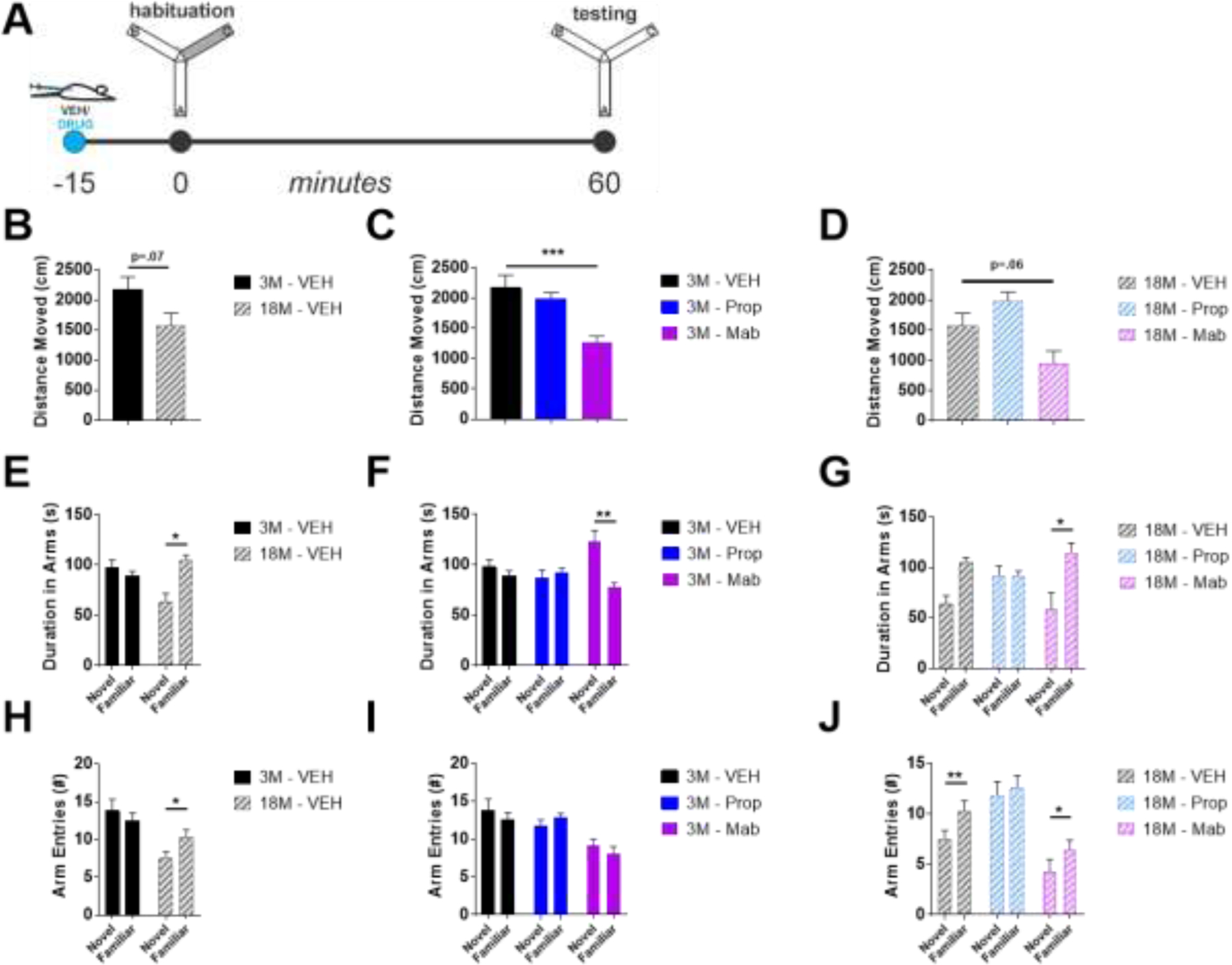
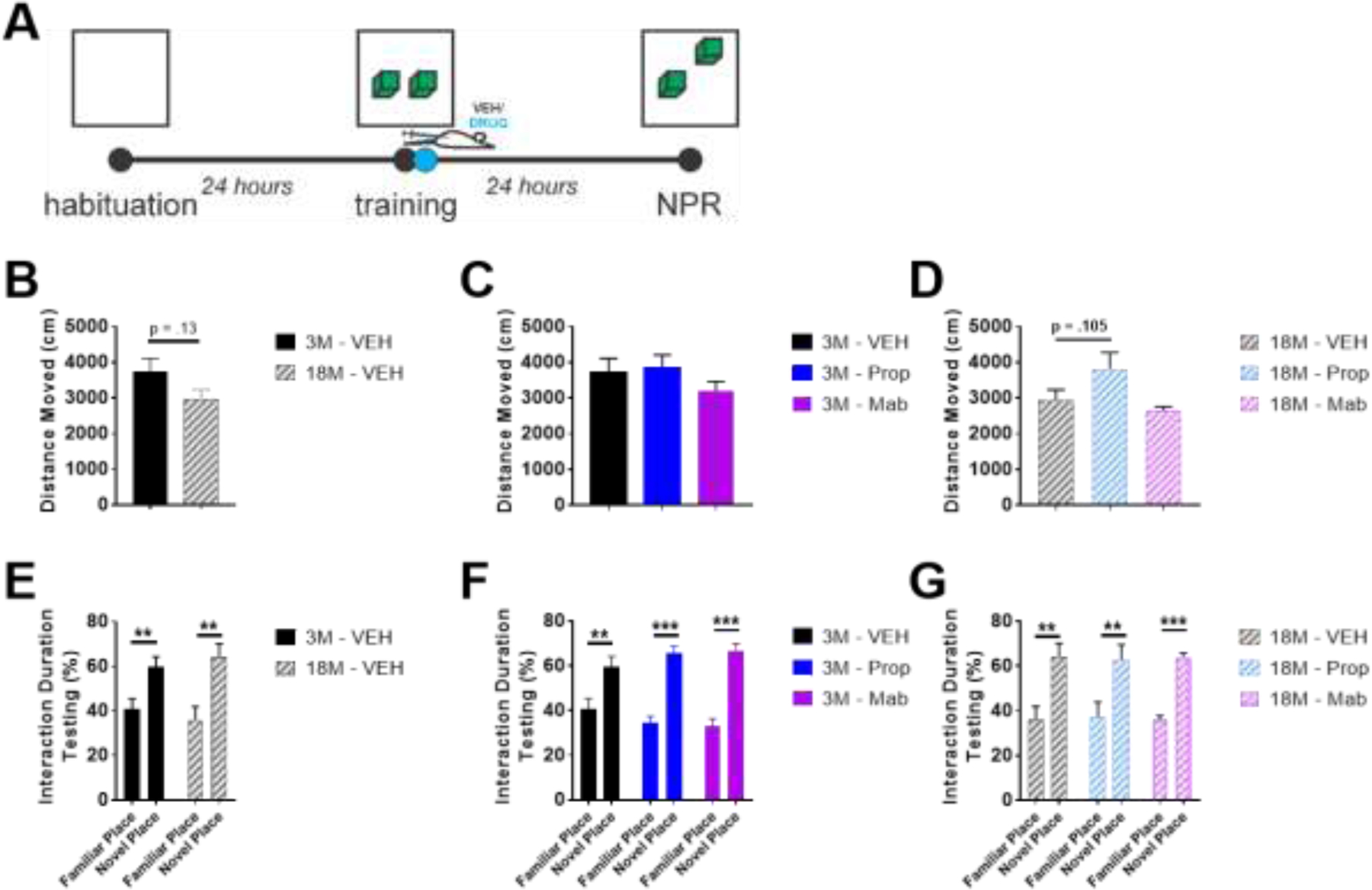
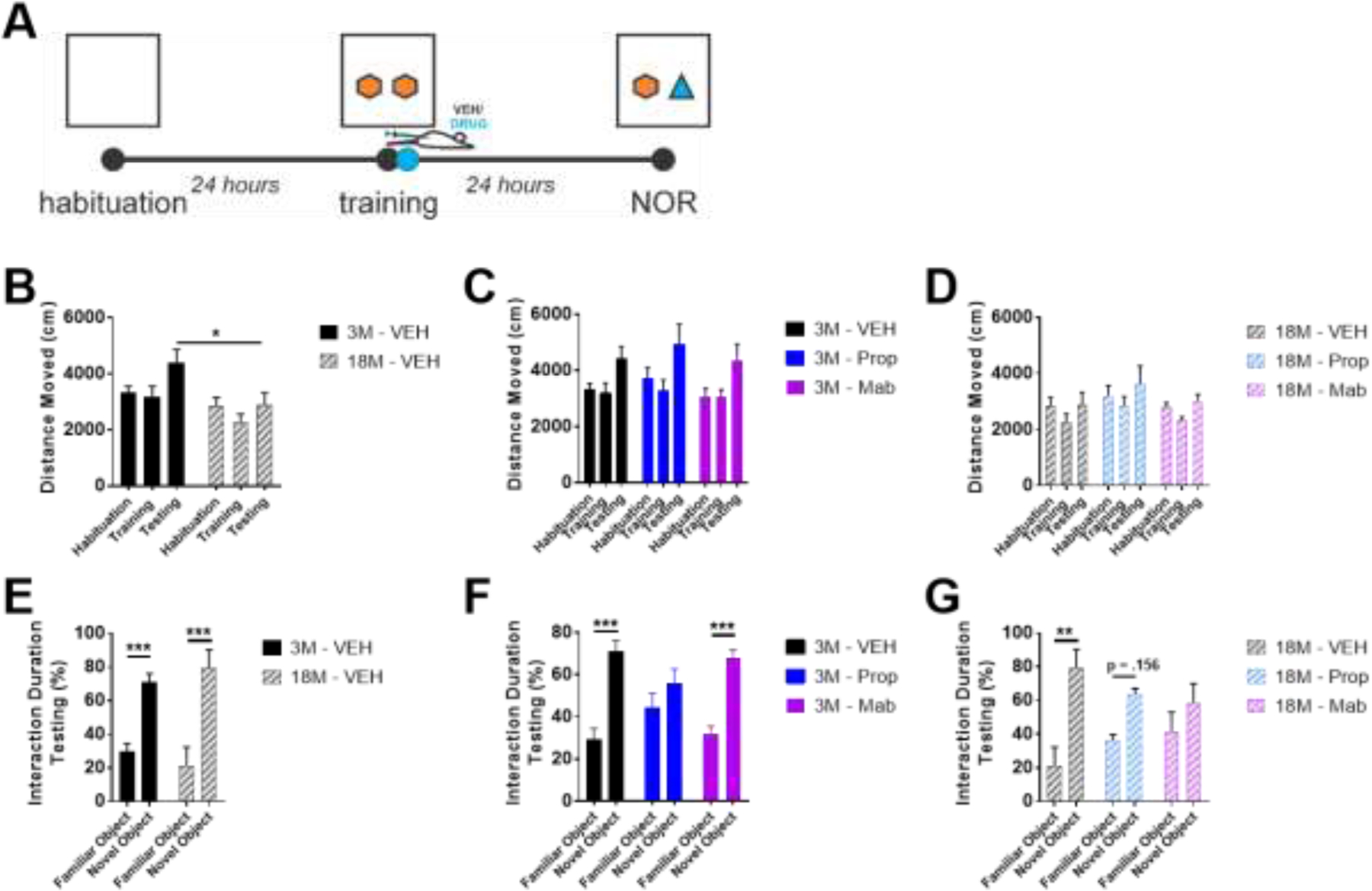
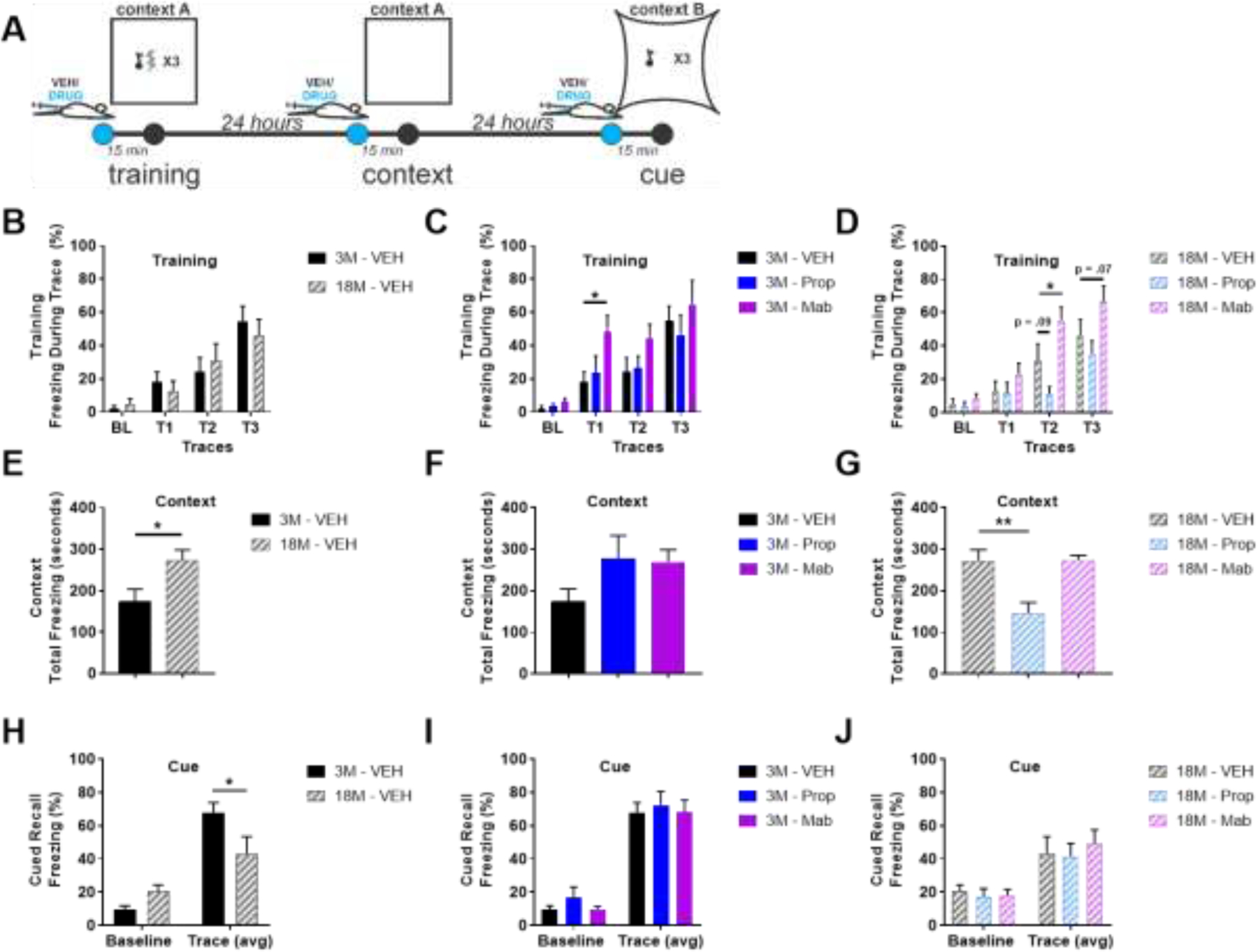
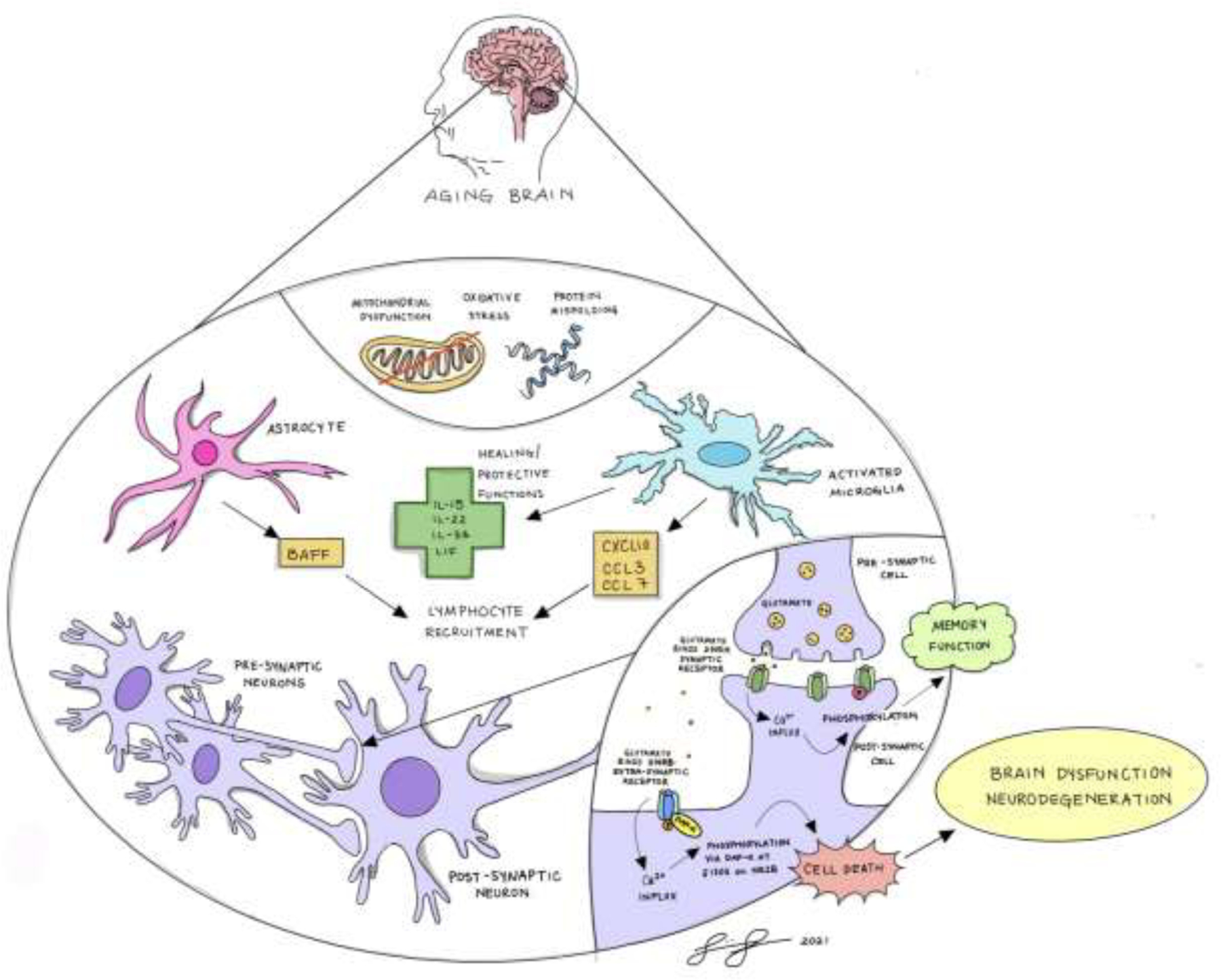
The locus coeruleus (LC) provides the primary noradrenergic input to the forebrain and hippocampus, and may be vulnerable to degeneration and contribute to age-related cognitive decline and neuroinflammation. Additionally, inhibition of noradrenergic transmission by brain-permeable beta-blockers could exacerbate cognitive impairment. This study examined effects of age and acute beta-blocker administration on LC and hippocampus pathology, neuroinflammation and learning and memory behavior in mice. Male mice, 3 and 18 months old, were administered propranolol (beta-blocker) or mabuterol (beta-adrenergic agonist) acutely around behavioral assessment. Terminal inflammatory markers in plasma, hippocampus and LC were assessed alongside histopathology. An increase in hippocampal and LC microgliosis and inflammatory proteins in the hippocampus was detected in aged mice. We report pathological hyperphosphorylation of the postsynaptic NMDA receptor subunit 2B (NR2B) in the hippocampus, suggesting neuronal hyperexcitability. Furthermore, the aged proteome revealed an induction in proteins related to energy metabolism, and mitochondria dysfunction in the LC and hippocampus. In a series of hippocampal dependent behavioral assessment tasks acute beta-adrenergic agonist or beta blocker administration altered learning and memory behavior in both aged and young mice. In Y-maze, propranolol and mabuterol differentially altered time spent in novel versus familiar arms in young and aged mice. Propranolol impaired Novel Object Recognition in both young and aged mice. Mabuterol enhanced trace learning in fear conditioning. Aged mice froze more to context and less to cue. Propranolol impaired contextual recall in aged mice. Concluding, aged mice show LC and hippocampus pathology and heightened effects of beta-adrenergic pharmacology on learning and memory.
Keywords: Aging; Behavior; Beta-blocker; Inflammation; Locus coeruleus; proteomics.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interest
None declared
Figures
Similar articles
-
Impact of high-fat diet on cognitive behavior and central and systemic inflammation with aging and sex differences in mice.Brain Behav Immun. 2024 May;118:334-354. doi: 10.1016/j.bbi.2024.02.025. Epub 2024 Feb 24. Brain Behav Immun. 2024. PMID: 38408498 Free PMC article.
-
Impact of noradrenergic inhibition on neuroinflammation and pathophysiology in mouse models of Alzheimer's disease.J Neuroinflammation. 2024 Dec 18;21(1):322. doi: 10.1186/s12974-024-03306-1. J Neuroinflammation. 2024. PMID: 39696597 Free PMC article.
-
Locus Coeruleus Ablation Exacerbates Cognitive Deficits, Neuropathology, and Lethality in P301S Tau Transgenic Mice.J Neurosci. 2018 Jan 3;38(1):74-92. doi: 10.1523/JNEUROSCI.1483-17.2017. Epub 2017 Nov 13. J Neurosci. 2018. PMID: 29133432 Free PMC article.
-
Multiple changes in noradrenergic mechanisms in the coeruleo-hippocampal pathway during aging. Structural and functional correlates in intraocular double grafts.Neurobiol Aging. 1989 Mar-Apr;10(2):117-24. doi: 10.1016/0197-4580(89)90020-1. Neurobiol Aging. 1989. PMID: 2542818 Review.
-
The Cholinergic System, the Adrenergic System and the Neuropathology of Alzheimer's Disease.Int J Mol Sci. 2021 Jan 28;22(3):1273. doi: 10.3390/ijms22031273. Int J Mol Sci. 2021. PMID: 33525357 Free PMC article. Review.
Cited by
-
Daily biofeedback to modulate heart rate oscillations affects structural volume in hippocampal subregions targeted by the locus coeruleus in older adults but not younger adults.Neurobiol Aging. 2023 Dec;132:85-99. doi: 10.1016/j.neurobiolaging.2023.08.010. Epub 2023 Sep 1. Neurobiol Aging. 2023. PMID: 37769491 Free PMC article. Clinical Trial.
-
Beta-2 adrenergic receptor agonism alters astrocyte phagocytic activity and has potential applications to psychiatric disease.Discov Ment Health. 2023 Nov 30;3(1):27. doi: 10.1007/s44192-023-00050-5. Discov Ment Health. 2023. PMID: 38036718 Free PMC article.
-
Possible role of locus coeruleus neuronal loss in age-related memory and attention deficits.Front Neurosci. 2023 Aug 24;17:1264253. doi: 10.3389/fnins.2023.1264253. eCollection 2023. Front Neurosci. 2023. PMID: 37694113 Free PMC article.
-
Locus Coeruleus and Noradrenergic Pharmacology in Neurodegenerative Disease.Handb Exp Pharmacol. 2024;285:555-616. doi: 10.1007/164_2023_677. Handb Exp Pharmacol. 2024. PMID: 37495851 Review.
-
Environmentally Toxic Solid Nanoparticles in Noradrenergic and Dopaminergic Nuclei and Cerebellum of Metropolitan Mexico City Children and Young Adults with Neural Quadruple Misfolded Protein Pathologies and High Exposures to Nano Particulate Matter.Toxics. 2022 Mar 29;10(4):164. doi: 10.3390/toxics10040164. Toxics. 2022. PMID: 35448425 Free PMC article.
References
-
- Amrhein V, et al., 2019. Scientists rise up against statistical significance. Nature. 567, 305–307. - PubMed
-
- Bialik S, Kimchi A, 2006. The death-associated protein kinases: structure, function, and beyond. Annu Rev Biochem. 75, 189–210. - PubMed
-
- Braak H, Del TK, 2011. The pathological process underlying Alzheimer’s disease in individuals under thirty. Acta Neuropathol. 121, 171–181. - PubMed
-
- Braak H, et al., 2011. Stages of the pathologic process in Alzheimer disease: age categories from 1 to 100 years. J. Neuropathol. Exp. Neurol. 70, 960–969. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
