

Mechanisms of BRCA1-BARD1 nucleosome recognition and ubiquitylation
- PMID: 34321665
- PMCID: PMC8680157
- DOI: 10.1038/s41586-021-03716-8
Mechanisms of BRCA1-BARD1 nucleosome recognition and ubiquitylation
Erratum in
-
Author Correction: Mechanisms of BRCA1-BARD1 nucleosome recognition and ubiquitylation.Nature. 2021 Sep;597(7875):E5. doi: 10.1038/s41586-021-03881-w. Nature. 2021. PMID: 34404953 No abstract available.
Abstract
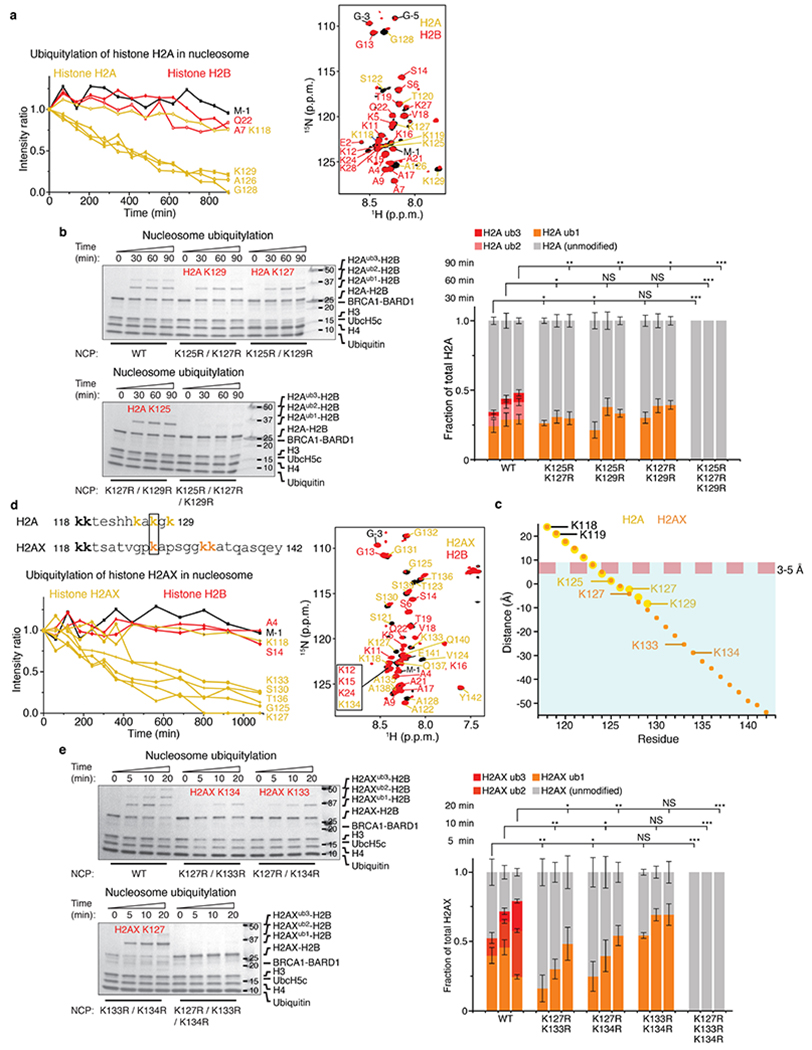
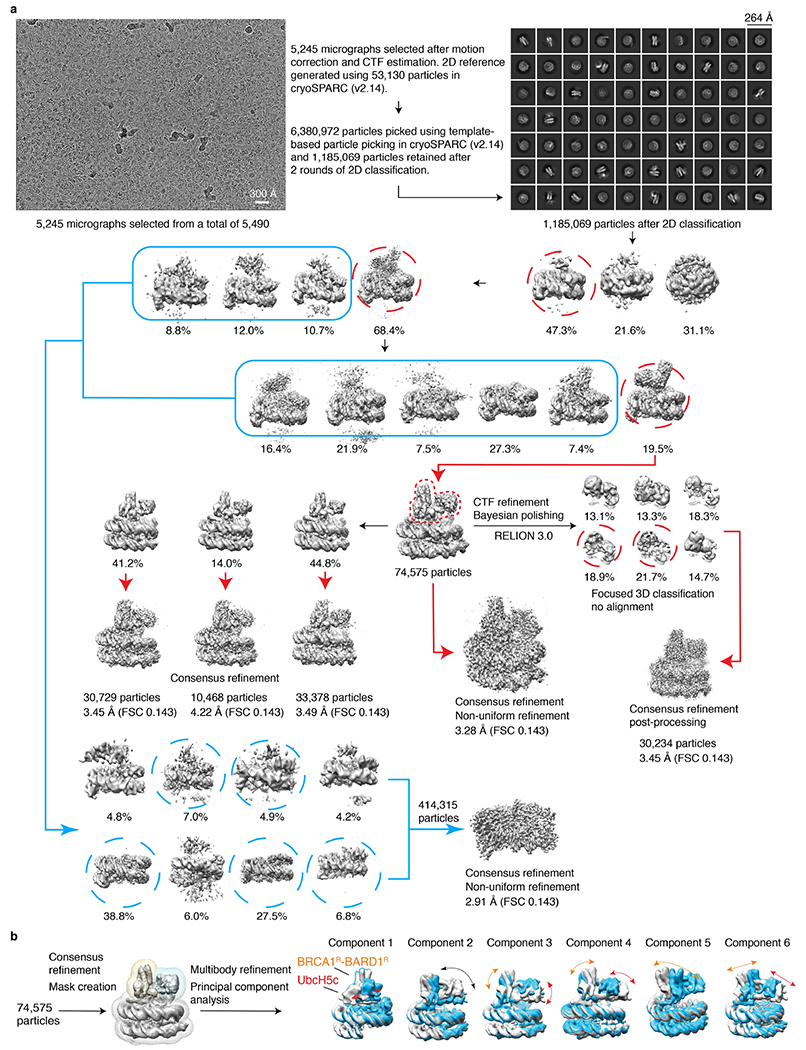
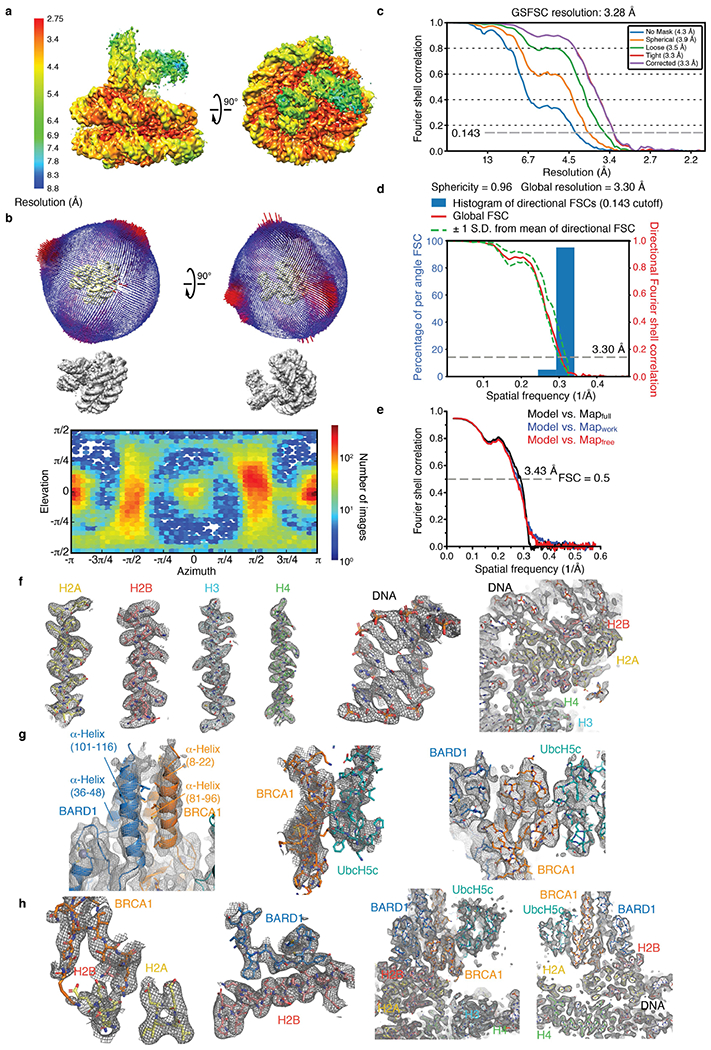
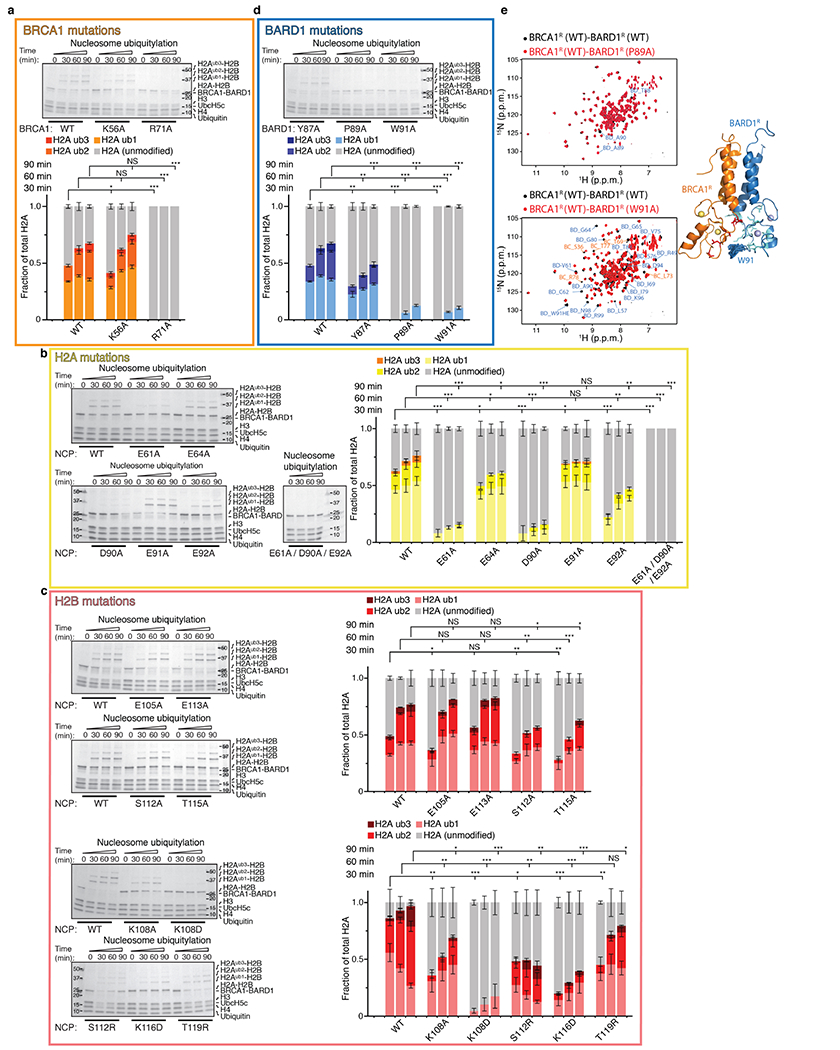
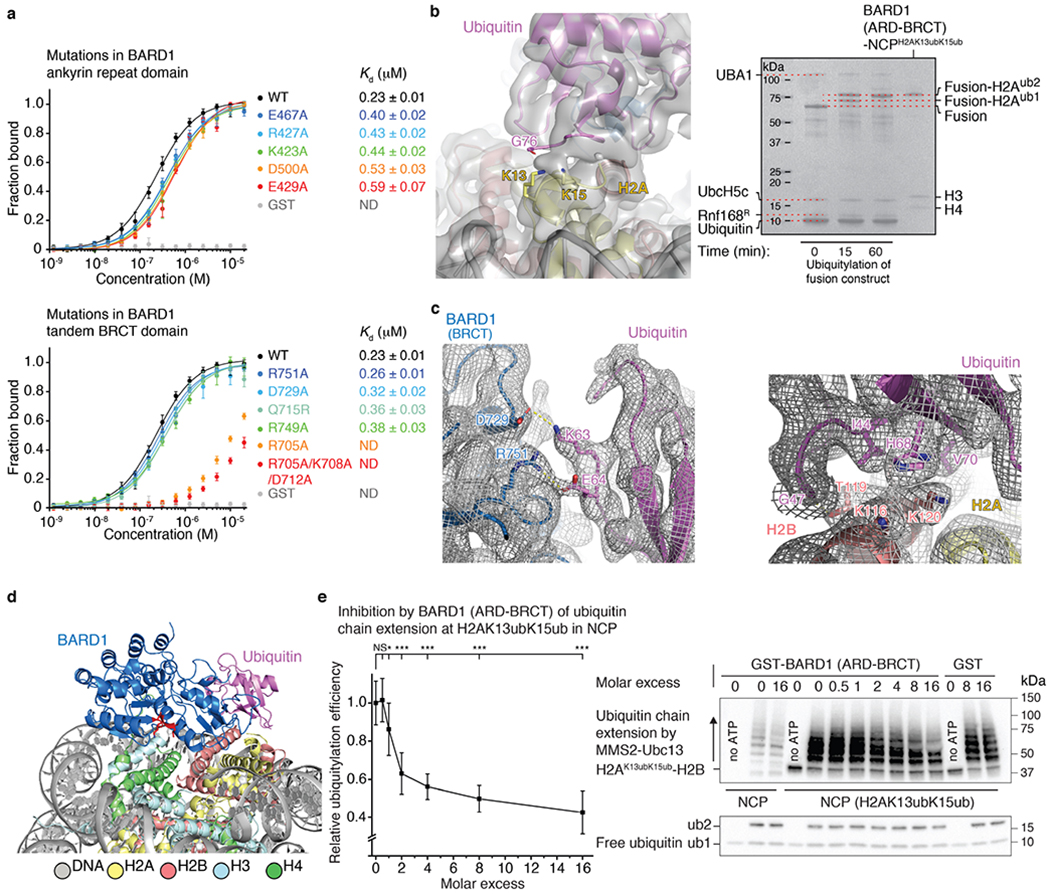
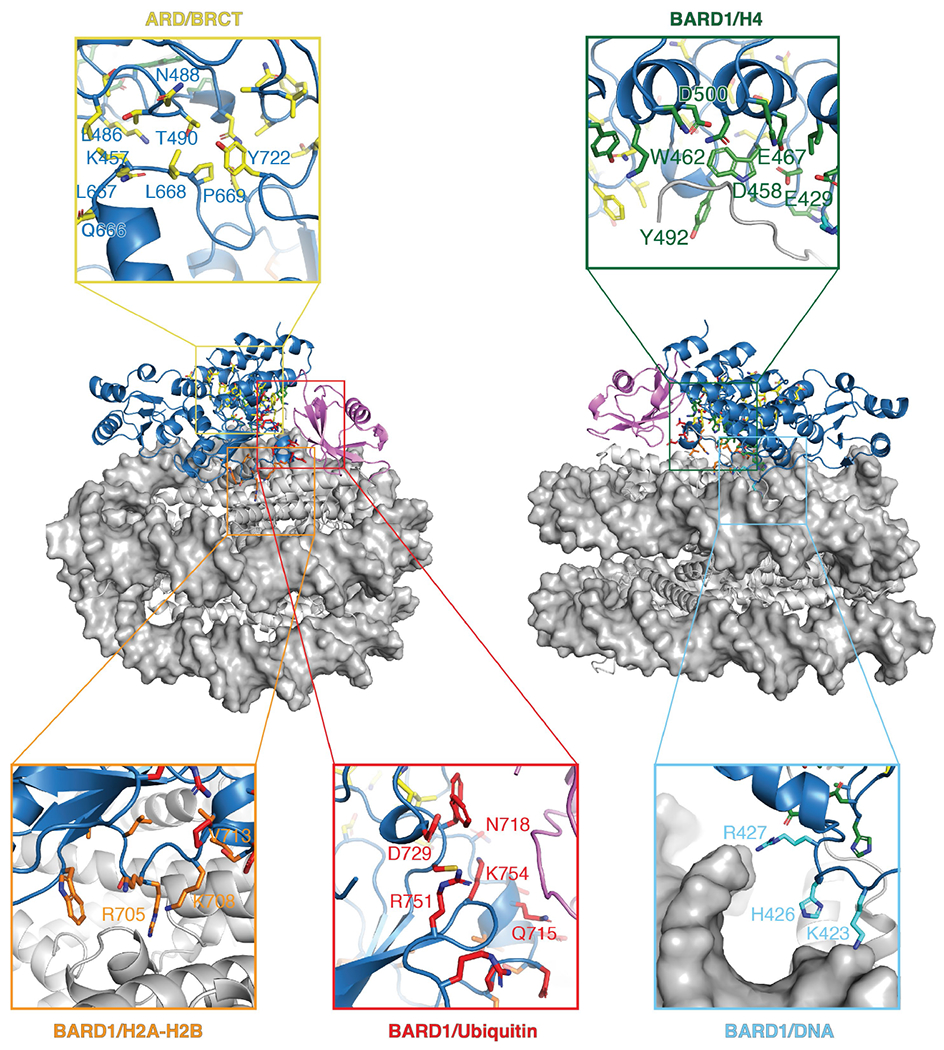
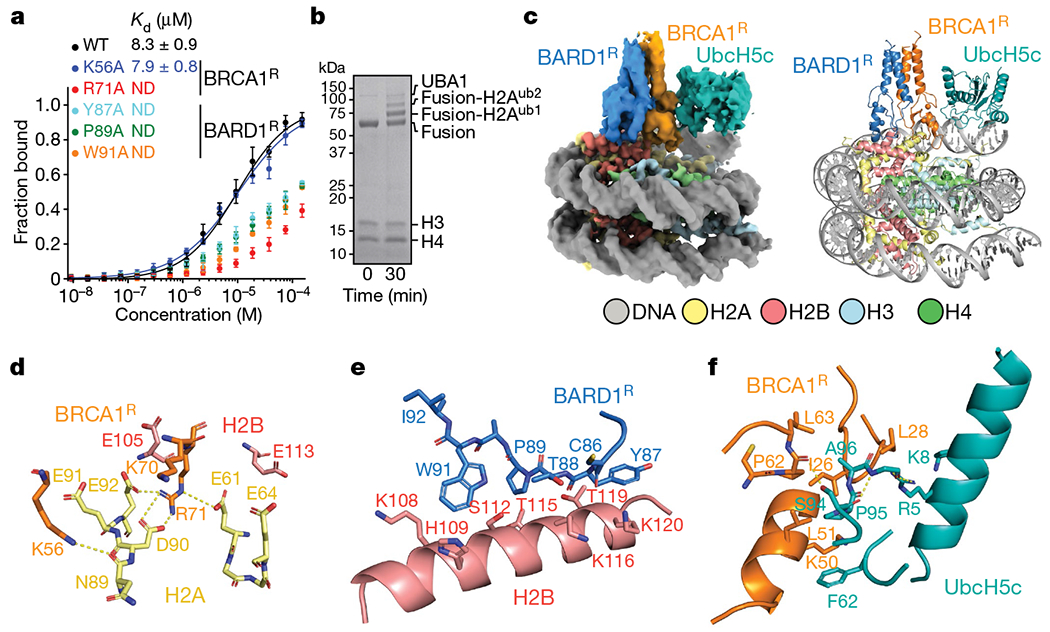
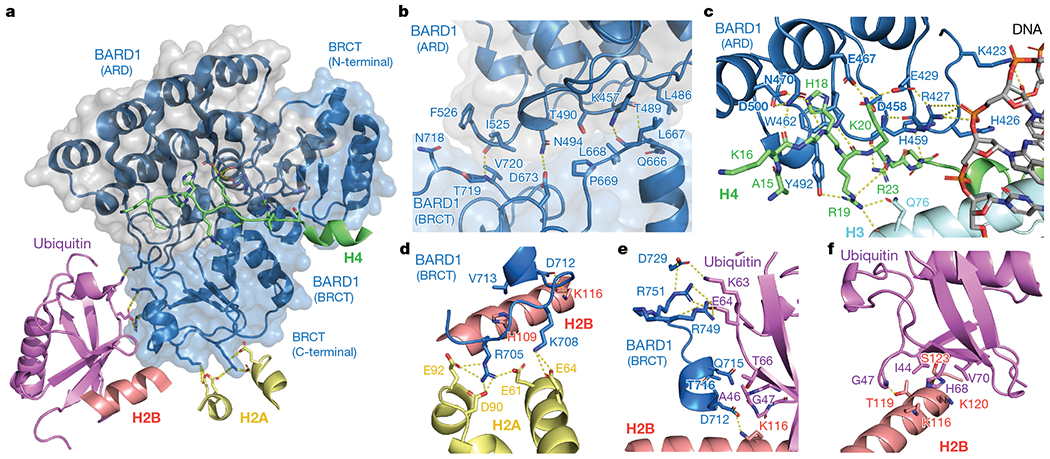
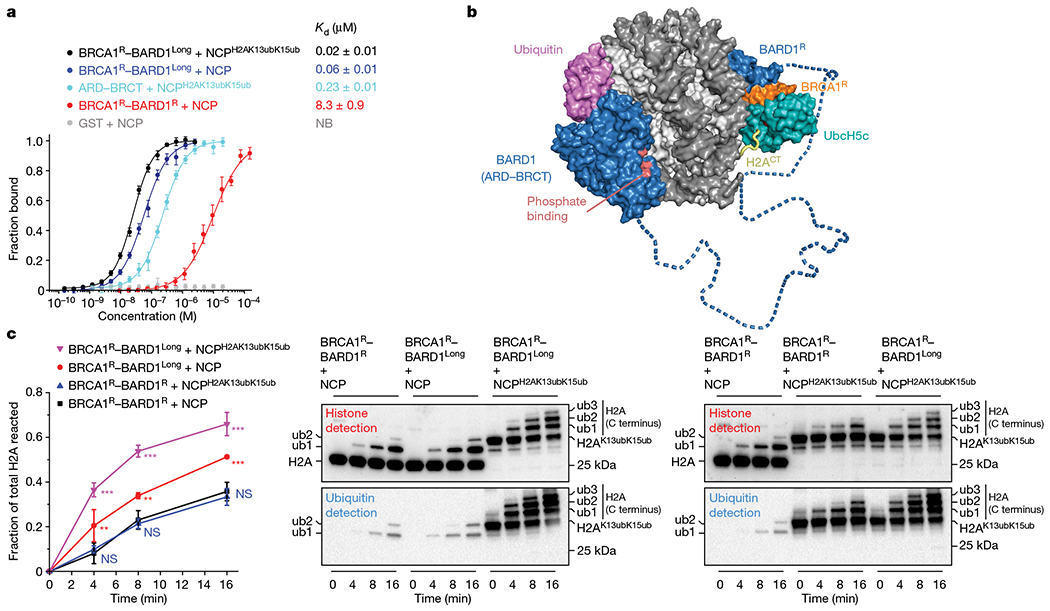
The BRCA1-BARD1 tumour suppressor is an E3 ubiquitin ligase necessary for the repair of DNA double-strand breaks by homologous recombination1-10. The BRCA1-BARD1 complex localizes to damaged chromatin after DNA replication and catalyses the ubiquitylation of histone H2A and other cellular targets11-14. The molecular bases for the recruitment to double-strand breaks and target recognition of BRCA1-BARD1 remain unknown. Here we use cryo-electron microscopy to show that the ankyrin repeat and tandem BRCT domains in BARD1 adopt a compact fold and bind to nucleosomal histones, DNA and monoubiquitin attached to H2A amino-terminal K13 or K15, two signals known to be specific for double-strand breaks15,16. We further show that RING domains17 in BRCA1-BARD1 orient an E2 ubiquitin-conjugating enzyme atop the nucleosome in a dynamic conformation, primed for ubiquitin transfer to the flexible carboxy-terminal tails of H2A and variant H2AX. Our work reveals a regulatory crosstalk in which recognition of monoubiquitin by BRCA1-BARD1 at the N terminus of H2A blocks the formation of polyubiquitin chains and cooperatively promotes ubiquitylation at the C terminus of H2A. These findings elucidate the mechanisms of BRCA1-BARD1 chromatin recruitment and ubiquitylation specificity, highlight key functions of BARD1 in both processes and explain how BRCA1-BARD1 promotes homologous recombination by opposing the DNA repair protein 53BP1 in post-replicative chromatin18-22. These data provide a structural framework to evaluate BARD1 variants and help to identify mutations that drive the development of cancer.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures
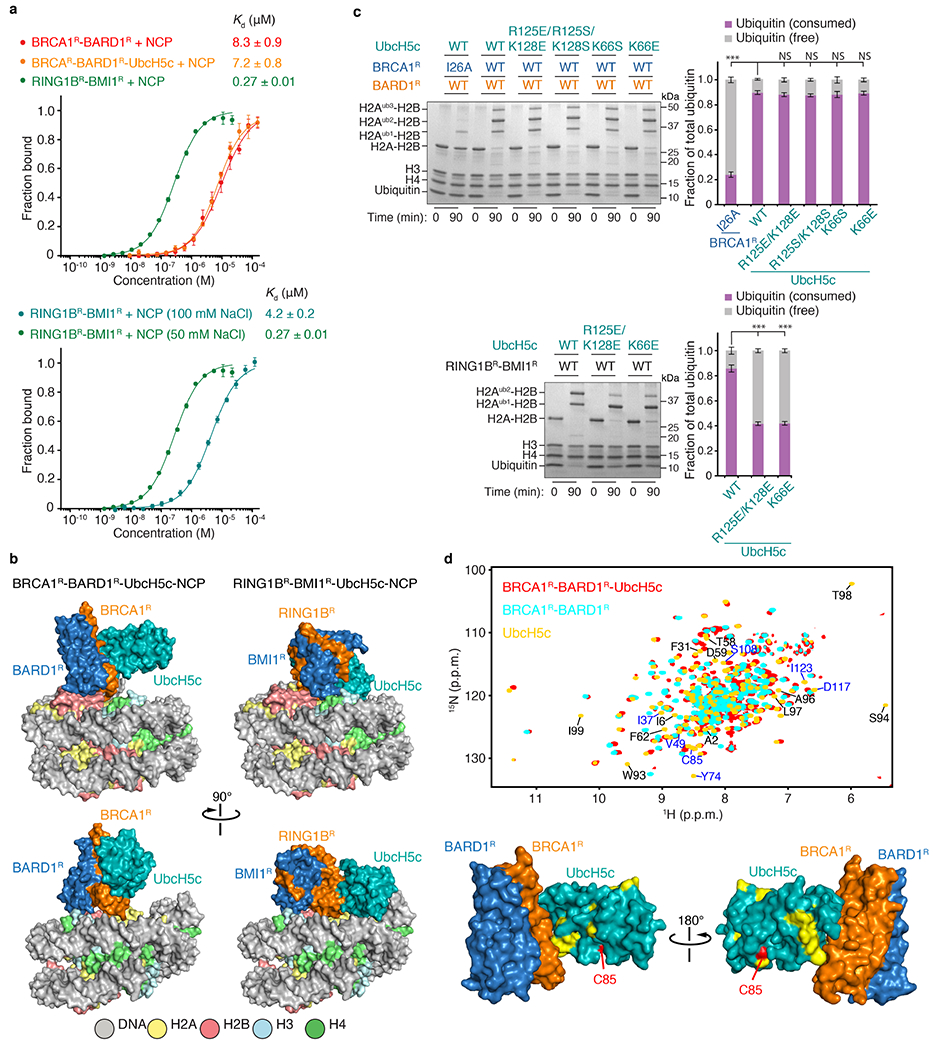
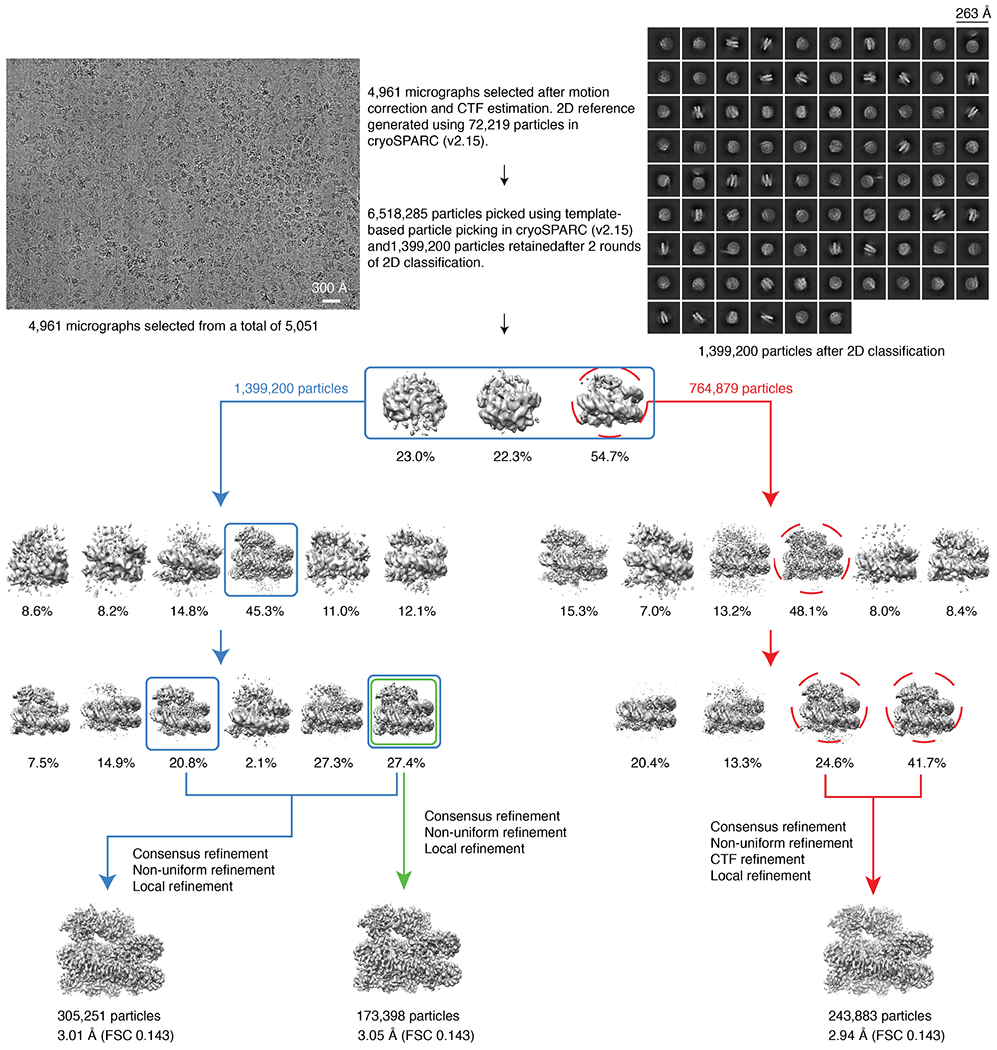
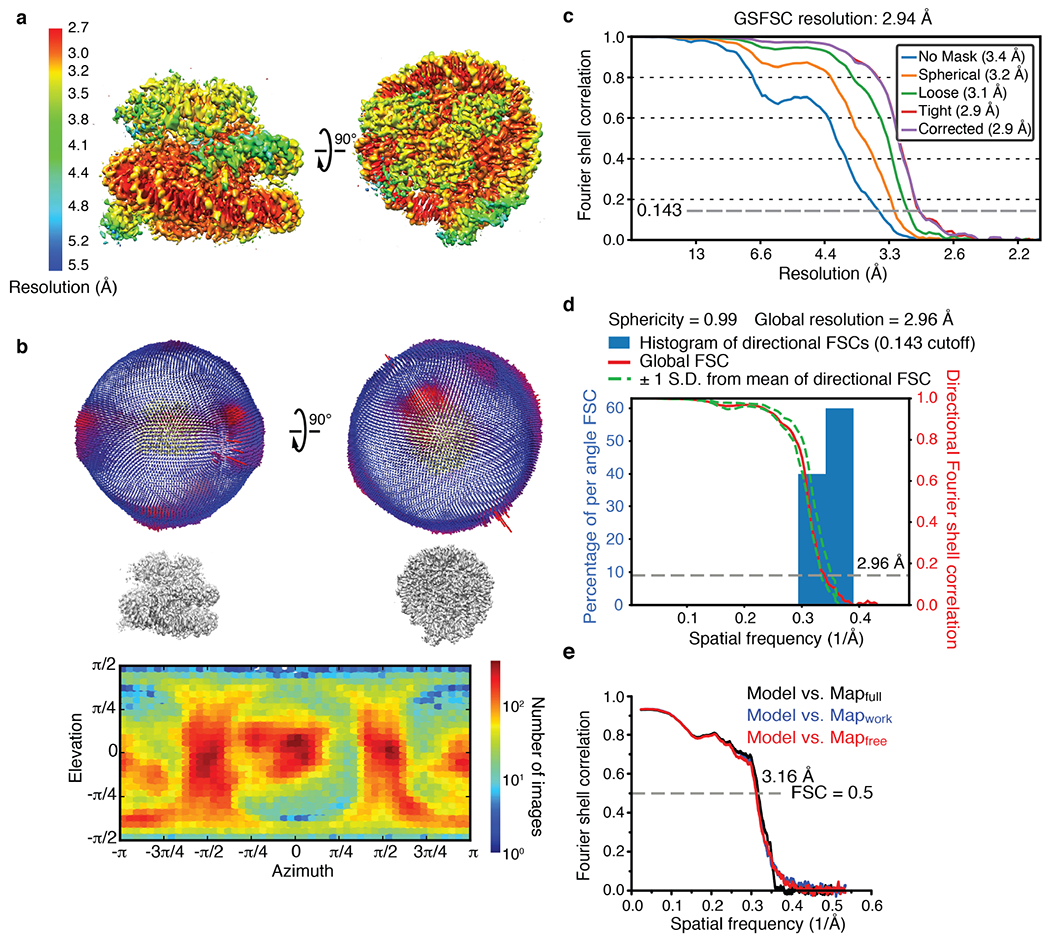
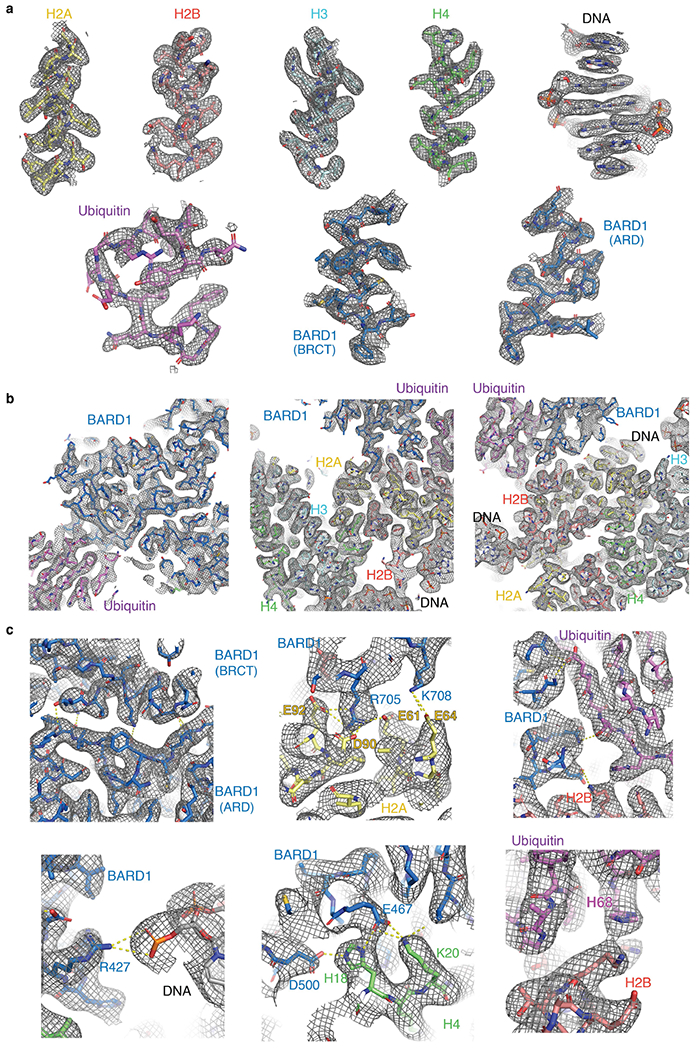

References
-
- Hall JM et al. Linkage of early-onset familial breast cancer to chromosome 17q21. Science 250, 1684–1689 (1990). - PubMed
-
- Miki Y et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science 266, 66–71 (1994). - PubMed
-
- Futreal PA et al. BRCA1 mutations in primary breast and ovarian carcinomas. Science 266, 120–122 (1994). - PubMed
-
- Friedman LS et al. Confirmation of BRCA1 by analysis of germline mutations linked to breast and ovarian cancer in ten families. Nat. Genet 8, 399–404 (1994). - PubMed
-
- Wu LC et al. Identification of a RING protein that can interact in vivo with the BRCA1 gene product. Nat. Genet 14, 430–440 (1996). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
