

Negative regulation of melatonin secretion by melatonin receptors in ovine pinealocytes
- PMID: 34324562
- PMCID: PMC8320996
- DOI: 10.1371/journal.pone.0255249
Negative regulation of melatonin secretion by melatonin receptors in ovine pinealocytes
Abstract
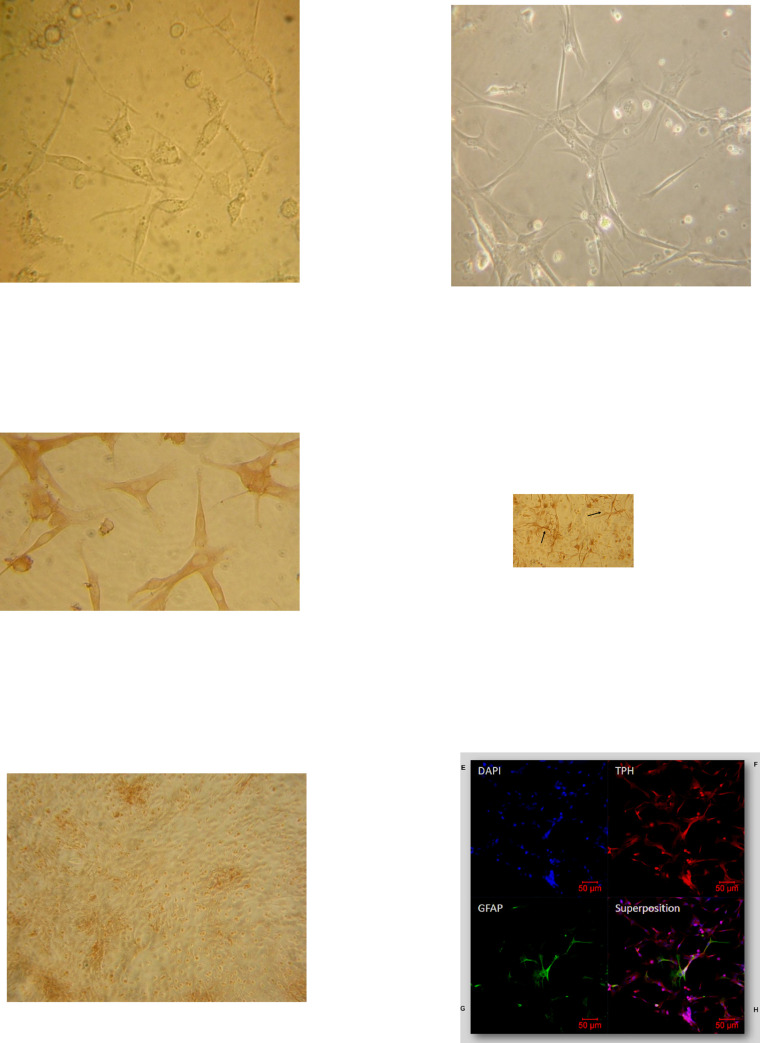
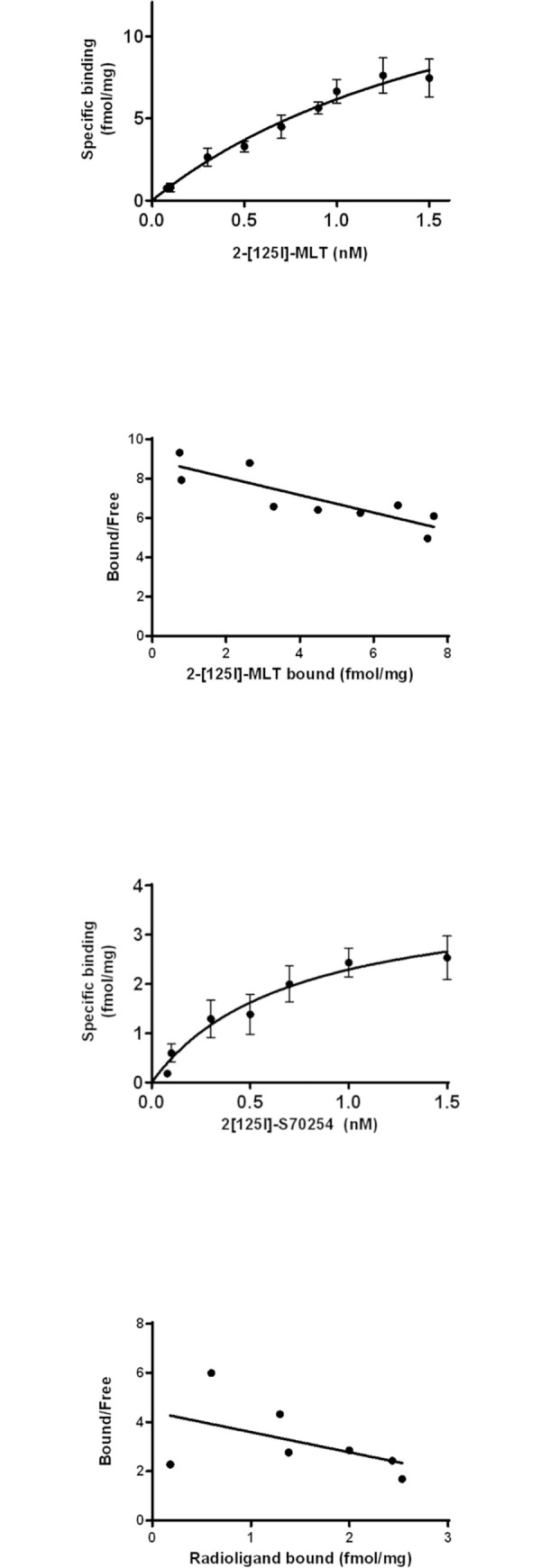
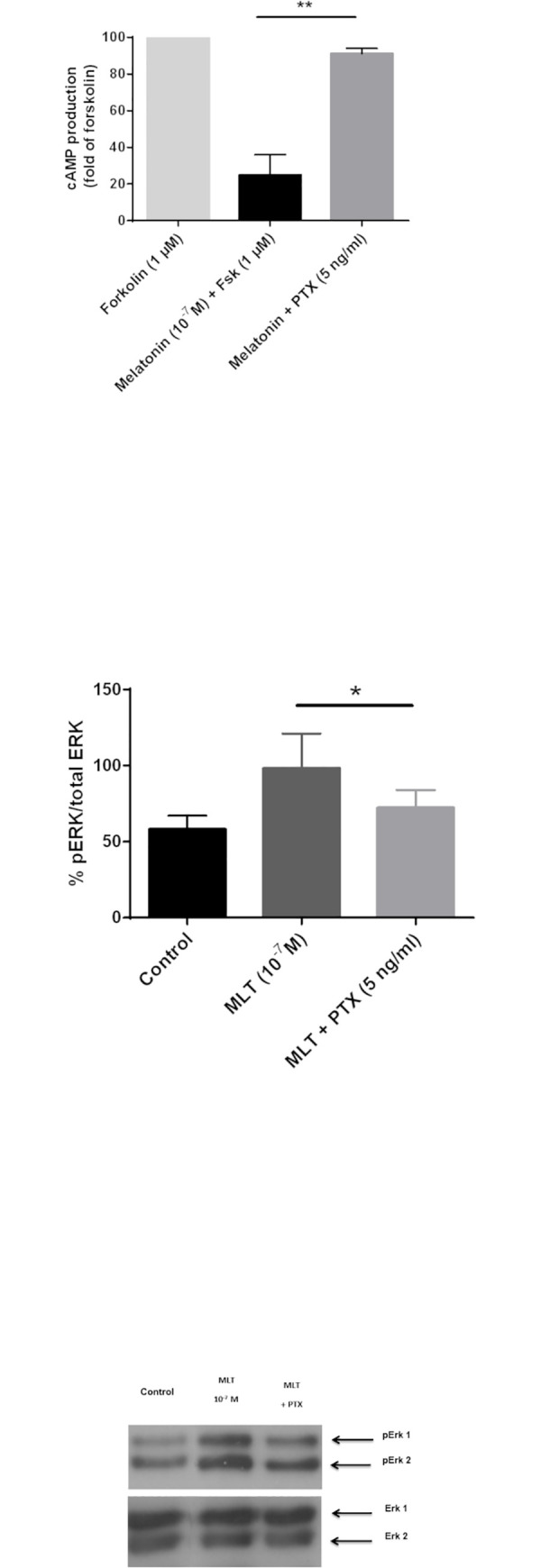
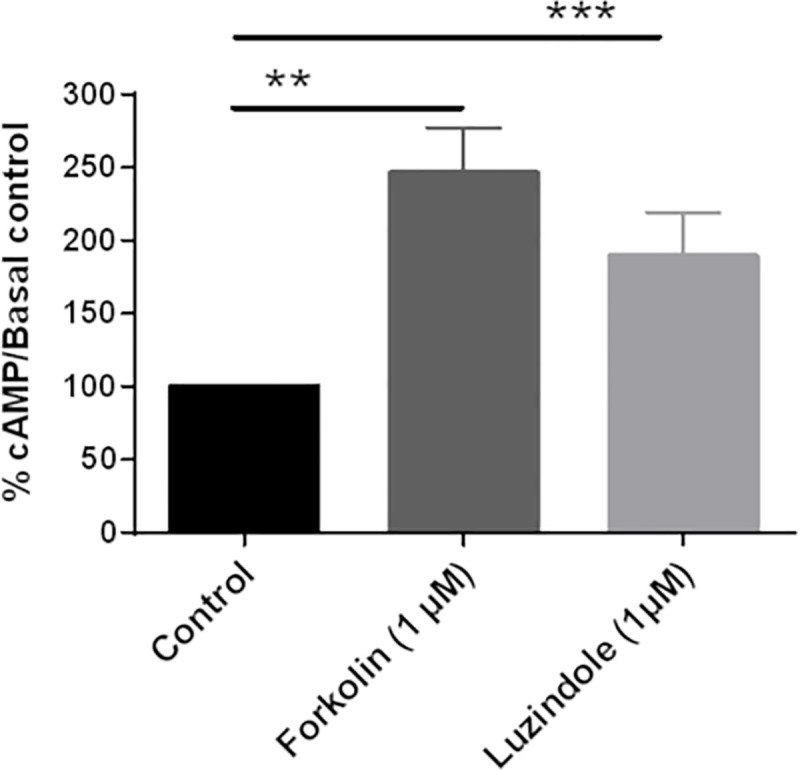
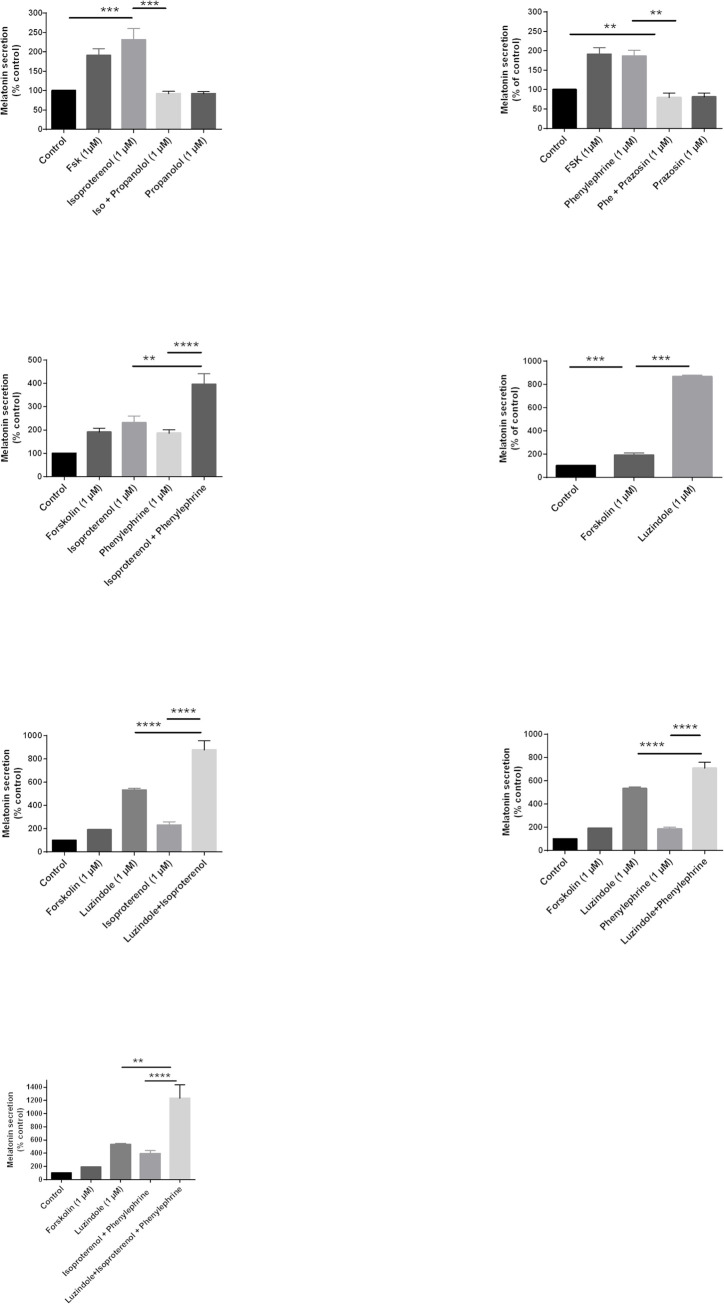
Melatonin (MLT) is a biological modulator of circadian and seasonal rhythms and reproduction. The photoperiodic information is detected by retinal photoreceptors and transmitted through nerve transmissions to the pineal gland, where MLT is synthesized and secreted at night into the blood. MLT interacts with two G protein-coupled receptors, MT1 and MT2. The aim of our work was to provide evidence for the presence of MLT receptors in the ovine pineal gland and define their involvement on melatonin secretion. For the first time, we identified the expression of MLT receptors with the specific 2-[125I]-MLT agonistic radioligand in ovin pinealocytes. The values of Kd and Bmax are 2.24 ± 1.1 nM and 20 ± 6.8 fmol/mg. MLT receptors are functional and inhibit cAMP production and activate ERK1/2 through pertussis toxin-sensitive Gi/o proteins. The MLT receptor antagonist/ inverse agonist luzindole increased cAMP production (189 ± 30%) and MLT secretion (866 ± 13%). The effect of luzindole on MLT secretion was additive with the effect of well-described activators of this pathway such as the β-adrenergic agonist isoproterenol and the α-adrenergic agonist phenylephrine. Co-incubation of all three compounds increased MLT secretion by 1236 ± 199%. These results suggest that MLT receptors are involved in the negative regulation of the synthesis of its own ligand in pinealocytes. While adrenergic receptors promote MLT secretion, MLT receptors mitigate this effect to limit the quantity of MLT secreted by the pineal gland.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Reiter RJ, Tan D, Terron MP, Flores LJ, Czarnocki Z. Melatonin and its metabolites: new findings regarding their production and their radical scavenging actions. Acta Biochim Pol. 2007;54(1):1‑9. - PubMed
-
- Shiu SYW, Pang B, Tam CW, Yao KM. Signal transduction of receptor-mediated antiproliferative action of melatonin on human prostate epithelial cells involves dual activation of Gαs and Gαq proteins: melatonin signaling in cell proliferation. J Pineal Res. 2010;49(3):301‑11. doi: 10.1111/j.1600-079X.2010.00795.x - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
