

Integration of transcriptional and metabolic control in macrophage activation
- PMID: 34328708
- PMCID: PMC8419687
- DOI: 10.15252/embr.202153251
Integration of transcriptional and metabolic control in macrophage activation
Abstract
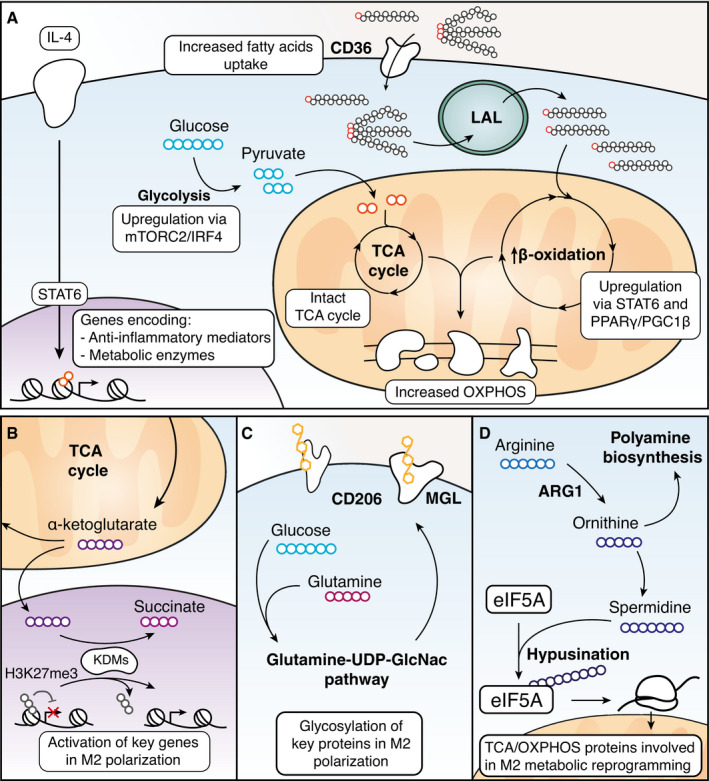
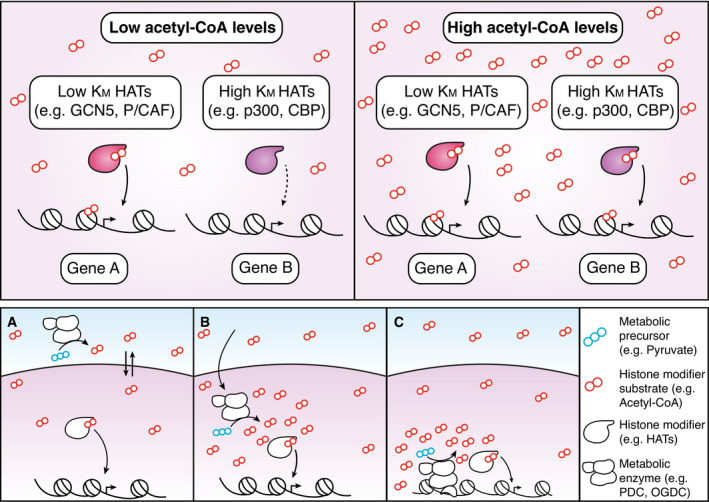
Macrophages react to microbial and endogenous danger signals by activating a broad panel of effector and homeostatic responses. Such responses entail rapid and stimulus-specific changes in gene expression programs accompanied by extensive rewiring of metabolism, with alterations in chromatin modifications providing one layer of integration of transcriptional and metabolic regulation. A systematic and mechanistic understanding of the mutual influences between signal-induced metabolic changes and gene expression is still lacking. Here, we discuss current evidence, controversies, knowledge gaps, and future areas of investigation on how metabolic and transcriptional changes are dynamically integrated during macrophage activation. The cross-talk between metabolism and inflammatory gene expression is in part accounted for by alterations in the production, usage, and availability of metabolic intermediates that impact the macrophage epigenome. In addition, stimulus-inducible gene expression changes alter the production of inflammatory mediators, such as nitric oxide, that in turn modulate the activity of metabolic enzymes thus determining complex regulatory loops. Critical issues remain to be understood, notably whether and how metabolic rewiring can bring about gene-specific (as opposed to global) expression changes.
Keywords: epigenetics; inflammation; macrophages; metabolism; transcription.
© 2021 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Amit I, Winter DR, Jung S (2016) The role of the local environment and epigenetics in shaping macrophage identity and their effect on tissue homeostasis. Nat Immunol 17: 18–25 - PubMed
-
- Anstey NM, Weinberg JB, Hassanali MY, Mwaikambo ED, Manyenga D, Misukonis MA, Arnelle DR, Hollis D, McDonald M, Granger DL (1996) Nitric oxide in Tanzanian children with malaria: inverse relationship between malaria severity and nitric oxide production/nitric oxide synthase type 2 expression. J Exp Med 184: 557–567 - PMC - PubMed
