

Novel mechanism for oscillations in catchbonded motor-filament complexes
- PMID: 34329628
- PMCID: PMC8510858
- DOI: 10.1016/j.bpj.2021.07.018
Novel mechanism for oscillations in catchbonded motor-filament complexes
Abstract
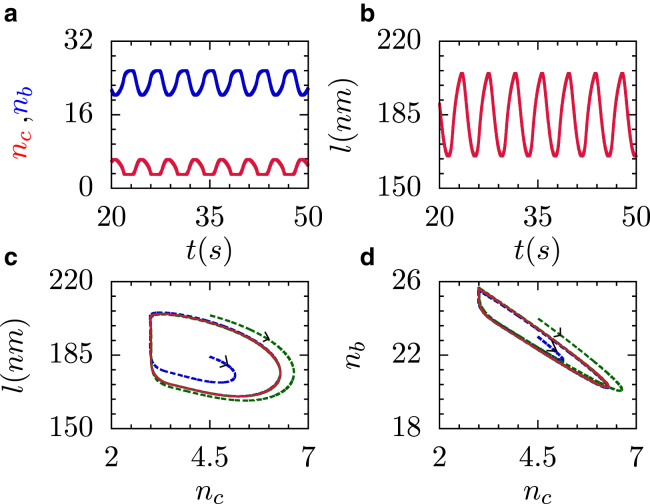
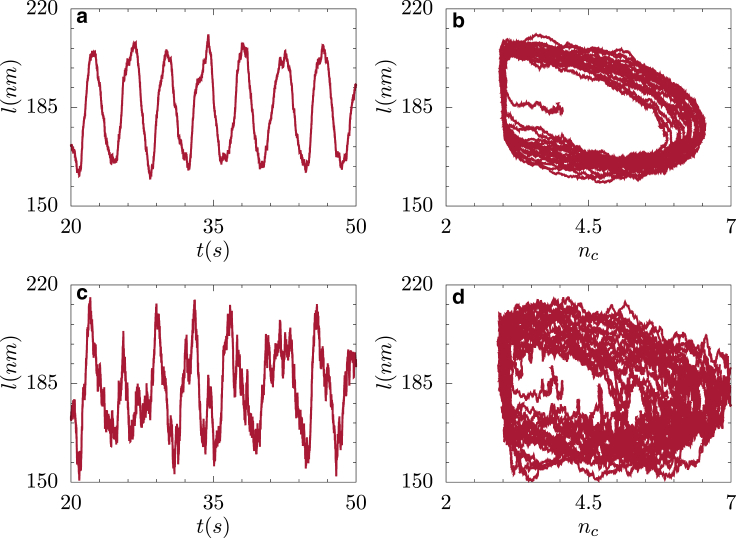
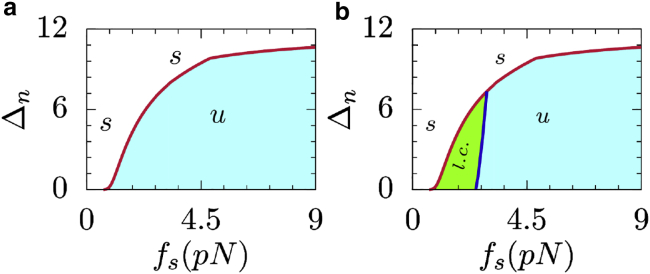
Generation of mechanical oscillations is ubiquitous to a wide variety of intracellular processes, ranging from activity of muscle fibers to oscillations of the mitotic spindle. The activity of motors plays a vital role in maintaining the integrity of the mitotic spindle structure and generating spontaneous oscillations. Although the structural features and properties of the individual motors are well characterized, their implications on the functional behavior of motor-filament complexes are more involved. We show that force-induced allosteric deformations in dynein, which result in catchbonding behavior, provide a generic mechanism to generate spontaneous oscillations in motor-cytoskeletal filament complexes. The resultant phase diagram of such motor-filament systems-characterized by force-induced allosteric deformations-exhibits bistability and sustained limit-cycle oscillations in biologically relevant regimes, such as for catchbonded dynein. The results reported here elucidate the central role of this mechanism in fashioning a distinctive stability behavior and oscillations in motor-filament complexes such as mitotic spindles.
Copyright © 2021 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Albertson D.G. Formation of the first cleavage spindle in nematode embryos. Dev. Biol. 1984;101:61–72. - PubMed
-
- Grill S.W., Kruse K., Jülicher F. Theory of mitotic spindle oscillations. Phys. Rev. Lett. 2005;94:108104. - PubMed
-
- Günther S., Kruse K. Spontaneous waves in muscle fibres. New J. Phys. 2007;9:417.
-
- Sasaki D., Fujita H., Ishiwata S. Auto-oscillations of skinned myocardium correlating with heartbeat. J. Muscle Res. Cell Motil. 2005;26:93–101. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
