

Affinity maturation of SARS-CoV-2 neutralizing antibodies confers potency, breadth, and resilience to viral escape mutations
- PMID: 34331873
- PMCID: PMC8323339
- DOI: 10.1016/j.immuni.2021.07.008
Affinity maturation of SARS-CoV-2 neutralizing antibodies confers potency, breadth, and resilience to viral escape mutations
Abstract
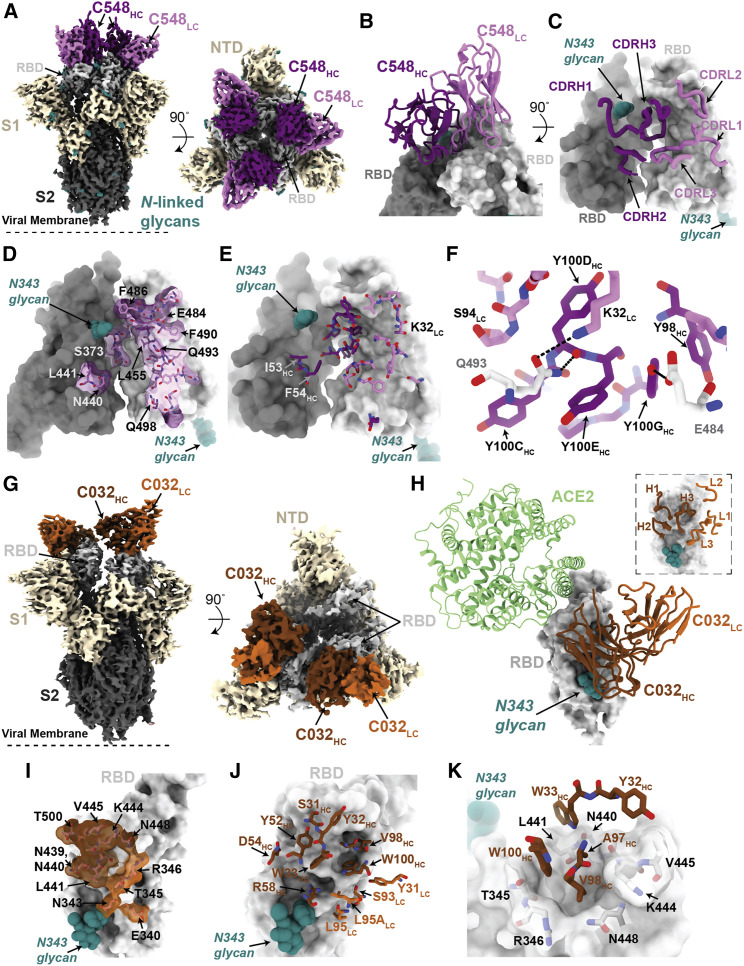
Antibodies elicited by infection accumulate somatic mutations in germinal centers that can increase affinity for cognate antigens. We analyzed 6 independent groups of clonally related severe acute respiratory syndrome-coronavirus-2 (SARS-CoV-2) Spike receptor-binding domain (RBD)-specific antibodies from 5 individuals shortly after infection and later in convalescence to determine the impact of maturation over months. In addition to increased affinity and neutralization potency, antibody evolution changed the mutational pathways for the acquisition of viral resistance and restricted neutralization escape options. For some antibodies, maturation imposed a requirement for multiple substitutions to enable escape. For certain antibodies, affinity maturation enabled the neutralization of circulating SARS-CoV-2 variants of concern and heterologous sarbecoviruses. Antibody-antigen structures revealed that these properties resulted from substitutions that allowed additional variability at the interface with the RBD. These findings suggest that increasing antibody diversity through prolonged or repeated antigen exposure may improve protection against diversifying SARS-CoV-2 populations, and perhaps against other pandemic threat coronaviruses.
Keywords: SARS-CoV-2; antibodies; neutralization.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The Rockefeller University has filed provisional patent applications in connection with this work on which M.C.N. (US patent 63/021,387) is listed as inventor. P.D.B. has served on an advisory board to Pfizer relating to SARS-CoV-2 vaccines.
Figures







Update of
-
Development of potency, breadth and resilience to viral escape mutations in SARS-CoV-2 neutralizing antibodies.bioRxiv [Preprint]. 2021 Mar 8:2021.03.07.434227. doi: 10.1101/2021.03.07.434227. bioRxiv. 2021. Update in: Immunity. 2021 Aug 10;54(8):1853-1868.e7. doi: 10.1016/j.immuni.2021.07.008. PMID: 33758864 Free PMC article. Updated. Preprint.
References
-
- Barnes C.O., West A.P., Jr., Huey-Tubman K.E., Hoffmann M.A.G., Sharaf N.G., Hoffman P.R., Koranda N., Gristick H.B., Gaebler C., Muecksch F., et al. Structures of Human Antibodies Bound to SARS-CoV-2 Spike Reveal Common Epitopes and Recurrent Features of Antibodies. Cell. 2020;182:828–842.e16. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
