

Oxidized GAPDH transfers S-glutathionylation to a nuclear protein Sirtuin-1 leading to apoptosis
- PMID: 34332079
- PMCID: PMC8432375
- DOI: 10.1016/j.freeradbiomed.2021.07.037
Oxidized GAPDH transfers S-glutathionylation to a nuclear protein Sirtuin-1 leading to apoptosis
Abstract
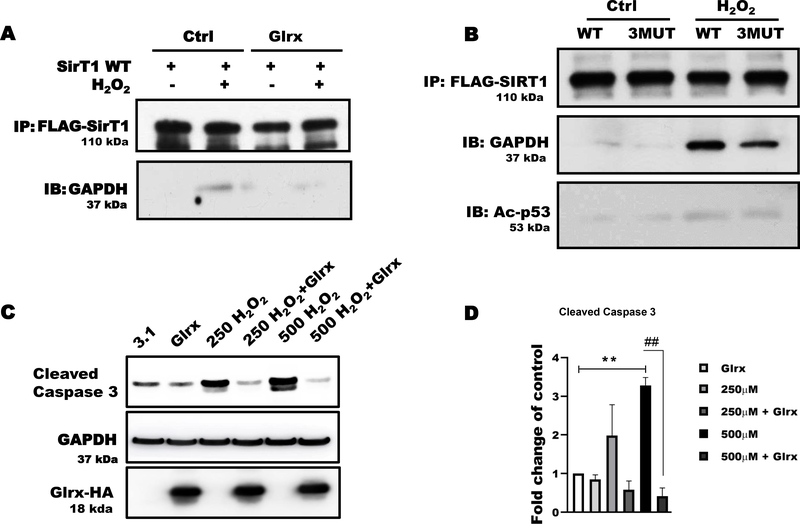
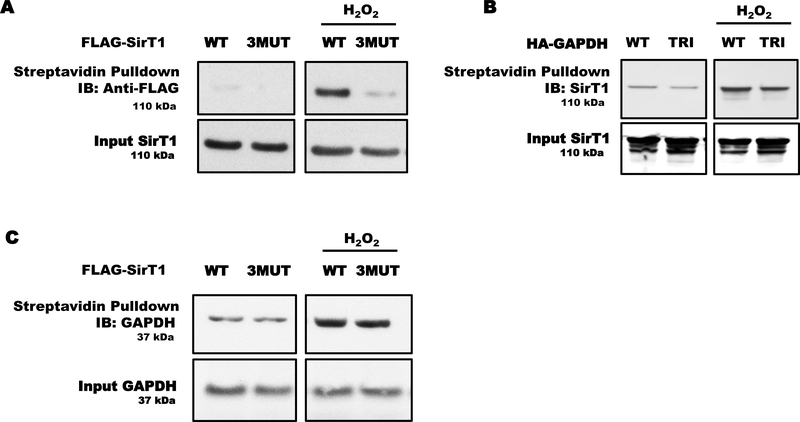
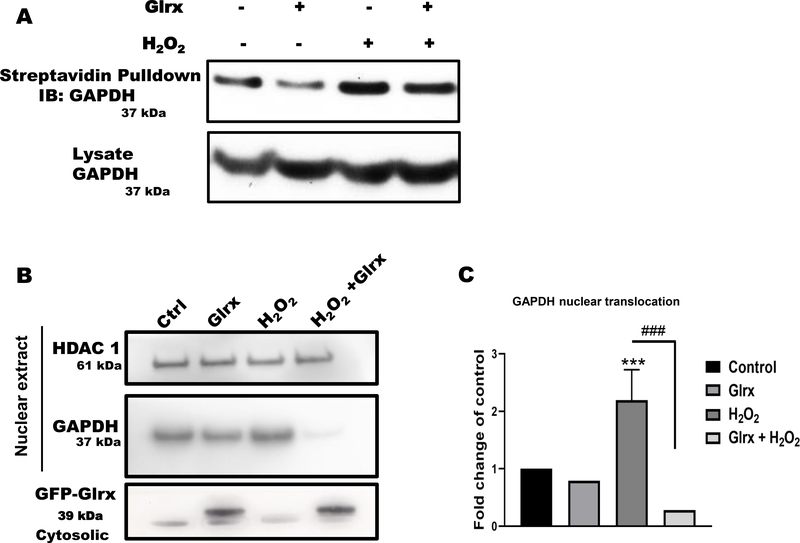
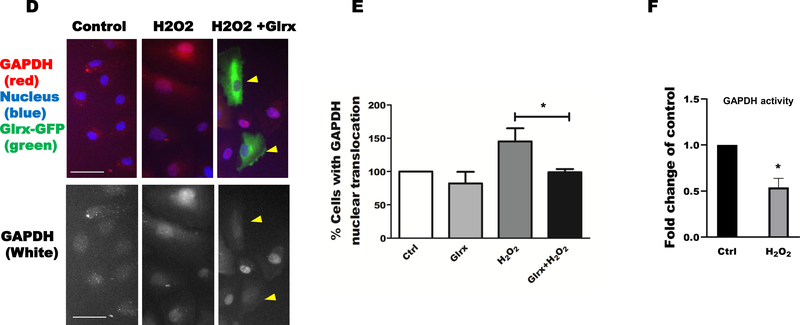
Aims: S-glutathionylation is a reversible oxidative modification of protein cysteines that plays a critical role in redox signaling. Glutaredoxin-1 (Glrx), a glutathione-specific thioltransferase, removes protein S-glutathionylation. Glrx, though a cytosolic protein, can activate a nuclear protein Sirtuin-1 (SirT1) by removing its S-glutathionylation. Glrx ablation causes metabolic abnormalities and promotes controlled cell death and fibrosis in mice. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), a key enzyme of glycolysis, is sensitive to oxidative modifications and involved in apoptotic signaling via the SirT1/p53 pathway in the nucleus. We aimed to elucidate the extent to which S-glutathionylation of GAPDH and glutaredoxin-1 contribute to GAPDH/SirT1/p53 apoptosis pathway.
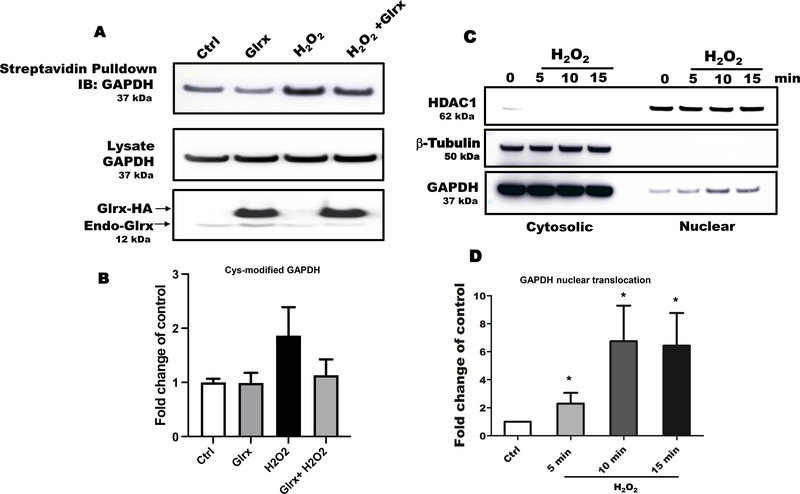
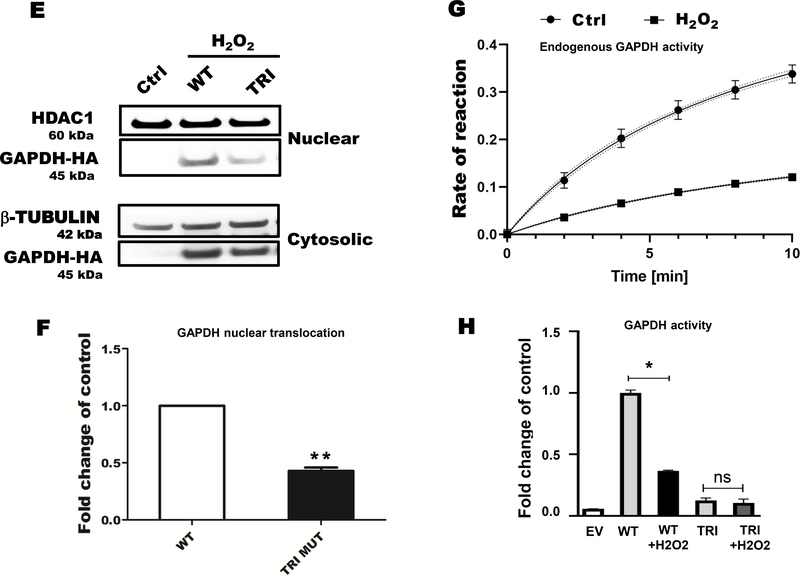
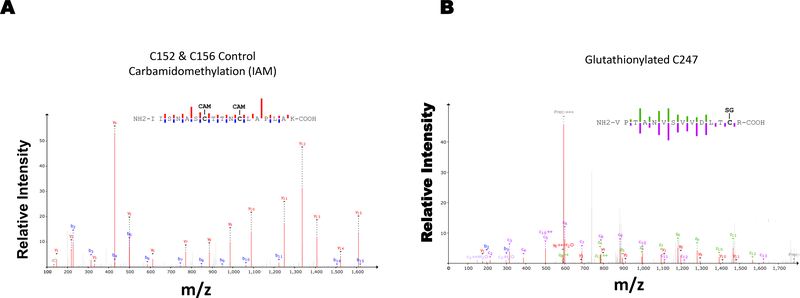
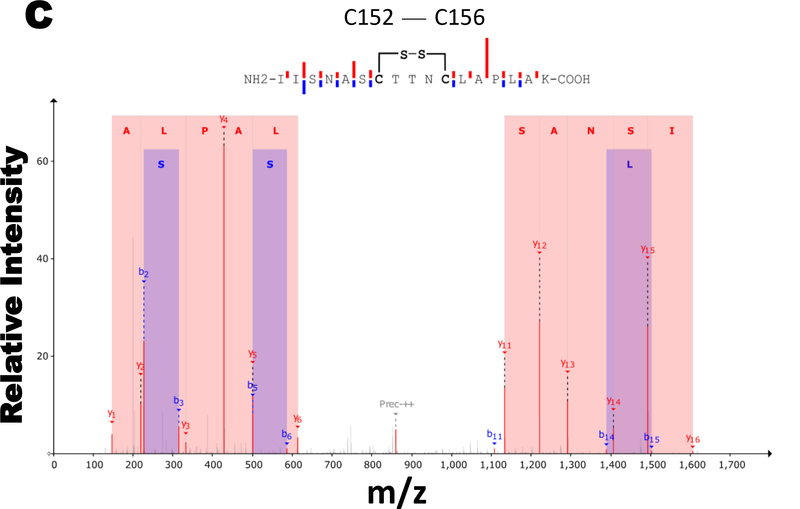
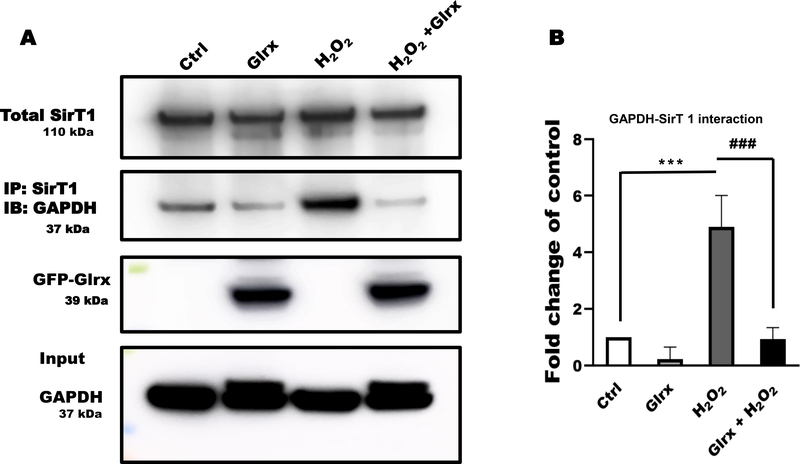
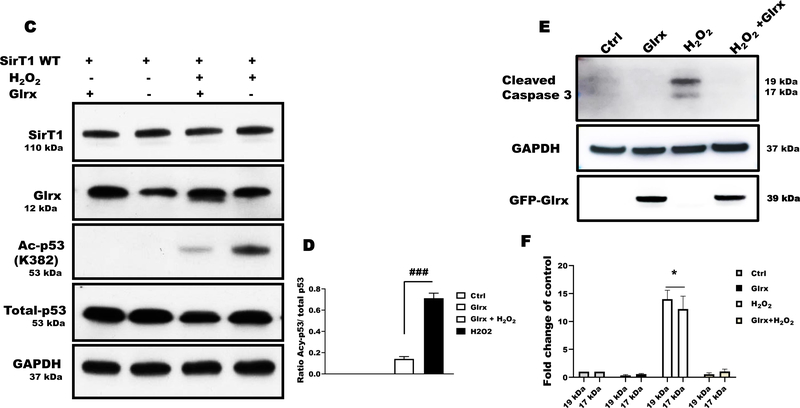
Results: Exposure of HEK 293T cells to hydrogen peroxide (H2O2) caused rapid S-glutathionylation and nuclear translocation of GAPDH. Nuclear GAPDH peaked 10-15 min after the addition of H2O2. Overexpression of Glrx or redox dead mutant GAPDH inhibited S-glutathionylation and nuclear translocation. Nuclear GAPDH formed a protein complex with SirT1 and exchanged S-glutathionylation to SirT1 and inhibited its deacetylase activity. Inactivated SirT1 remained stably bound to acetylated-p53 and initiated apoptotic signaling resulting in cleavage of caspase-3. We observed similar effects in human primary aortic endothelial cells suggesting the GAPDH/SirT1/p53 pathway as a common apoptotic mechanism.
Conclusions: Abundant GAPDH with its highly reactive-cysteine thiolate may function as a cytoplasmic rheostat to sense oxidative stress. S-glutathionylation of GAPDH may relay the signal to the nucleus where GAPDH trans-glutathionylates nuclear proteins such as SirT1 to initiate apoptosis. Glrx reverses GAPDH S-glutathionylation and prevents its nuclear translocation and cytoplasmic-nuclear redox signaling leading to apoptosis. Our data suggest that trans-glutathionylation is a critical step in apoptotic signaling and a potential mechanism that cytosolic Glrx controls nuclear transcription factors.
Keywords: GAPDH; Glutaredoxin; S-Glutathionylation; SirT1; Trans-Glutathionylation.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Redox Regulation via Glutaredoxin-1 and Protein S-Glutathionylation.Antioxid Redox Signal. 2020 Apr 1;32(10):677-700. doi: 10.1089/ars.2019.7963. Epub 2020 Jan 23. Antioxid Redox Signal. 2020. PMID: 31813265 Free PMC article. Review.
-
Redox Dysregulation of Vascular Smooth Muscle Sirtuin-1 in Thoracic Aortic Aneurysm in Marfan Syndrome.Arterioscler Thromb Vasc Biol. 2023 Aug;43(8):e339-e357. doi: 10.1161/ATVBAHA.123.319145. Epub 2023 Jun 8. Arterioscler Thromb Vasc Biol. 2023. PMID: 37288573 Free PMC article.
-
A redox-resistant sirtuin-1 mutant protects against hepatic metabolic and oxidant stress.J Biol Chem. 2014 Mar 14;289(11):7293-306. doi: 10.1074/jbc.M113.520403. Epub 2014 Jan 22. J Biol Chem. 2014. PMID: 24451382 Free PMC article.
-
Glutaredoxin-1 Deficiency Causes Fatty Liver and Dyslipidemia by Inhibiting Sirtuin-1.Antioxid Redox Signal. 2017 Aug 20;27(6):313-327. doi: 10.1089/ars.2016.6716. Epub 2017 Feb 16. Antioxid Redox Signal. 2017. PMID: 27958883 Free PMC article.
-
Regulation of neovascularization by S-glutathionylation via the Wnt5a/sFlt-1 pathway.Biochem Soc Trans. 2014 Dec;42(6):1665-70. doi: 10.1042/BST20140213. Biochem Soc Trans. 2014. PMID: 25399587 Free PMC article. Review.
Cited by
-
Origin of Elevated S-Glutathionylated GAPDH in Chronic Neurodegenerative Diseases.Int J Mol Sci. 2023 Mar 14;24(6):5529. doi: 10.3390/ijms24065529. Int J Mol Sci. 2023. PMID: 36982600 Free PMC article.
-
Modification of Glyceraldehyde-3-Phosphate Dehydrogenase with Nitric Oxide: Role in Signal Transduction and Development of Apoptosis.Biomolecules. 2021 Nov 8;11(11):1656. doi: 10.3390/biom11111656. Biomolecules. 2021. PMID: 34827652 Free PMC article. Review.
-
GAPDH: unveiling its impact as a key hypoxia-related player in head and neck squamous cell carcinoma tumor progression, prognosis, and therapeutic potential.Am J Cancer Res. 2023 Dec 15;13(12):5846-5867. eCollection 2023. Am J Cancer Res. 2023. PMID: 38187058 Free PMC article.
-
Role of Posttranslational Modifications of Proteins in Cardiovascular Disease.Oxid Med Cell Longev. 2022 Jul 9;2022:3137329. doi: 10.1155/2022/3137329. eCollection 2022. Oxid Med Cell Longev. 2022. PMID: 35855865 Free PMC article. Review.
-
Sirtuins at the Crossroads between Mitochondrial Quality Control and Neurodegenerative Diseases: Structure, Regulation, Modifications, and Modulators.Aging Dis. 2023 Jun 1;14(3):794-824. doi: 10.14336/AD.2022.1123. Aging Dis. 2023. PMID: 37191431 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
