

The osteocyte as a signaling cell
- PMID: 34337974
- PMCID: PMC8858675
- DOI: 10.1152/physrev.00043.2020
The osteocyte as a signaling cell
Abstract
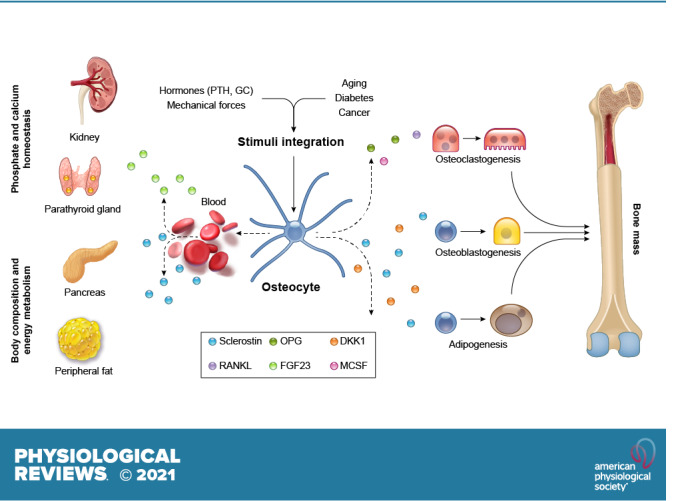
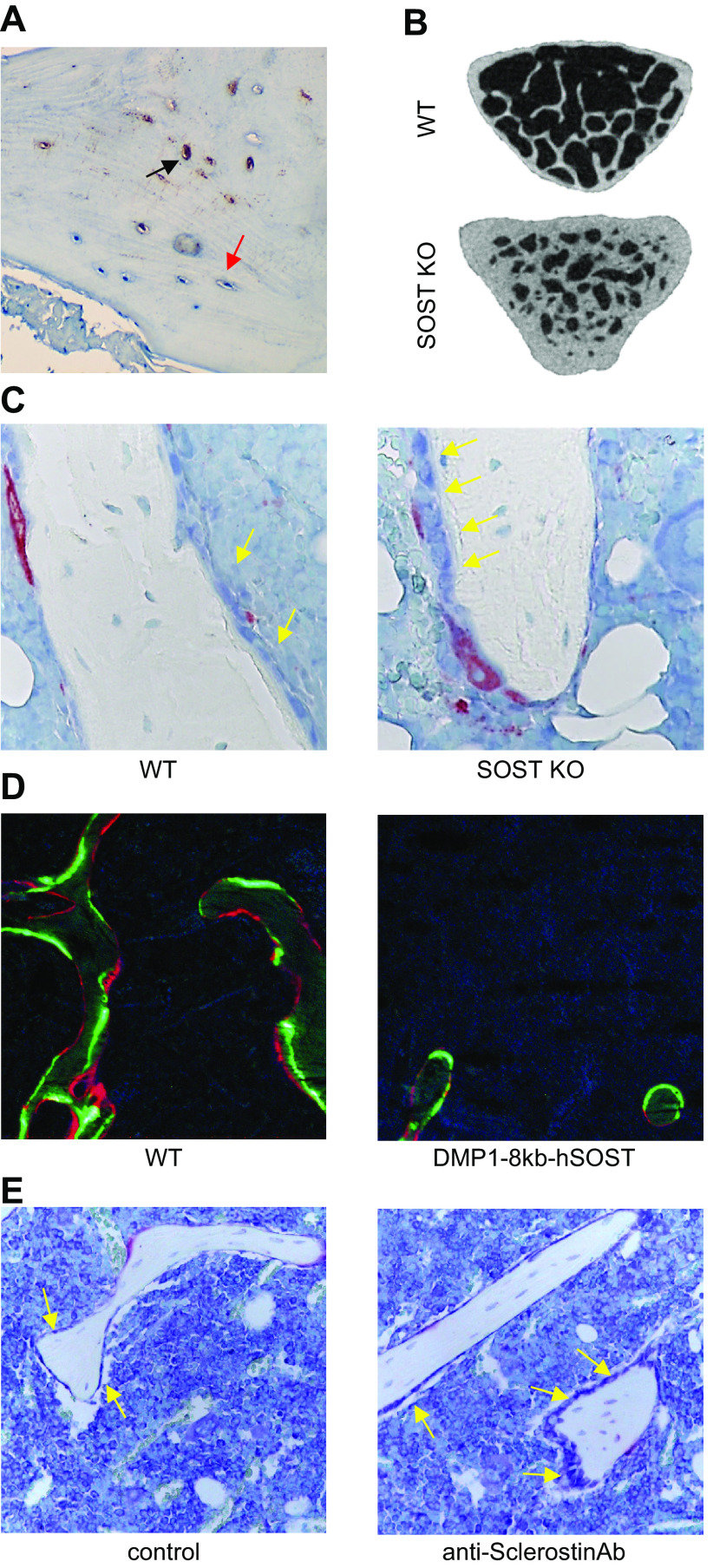
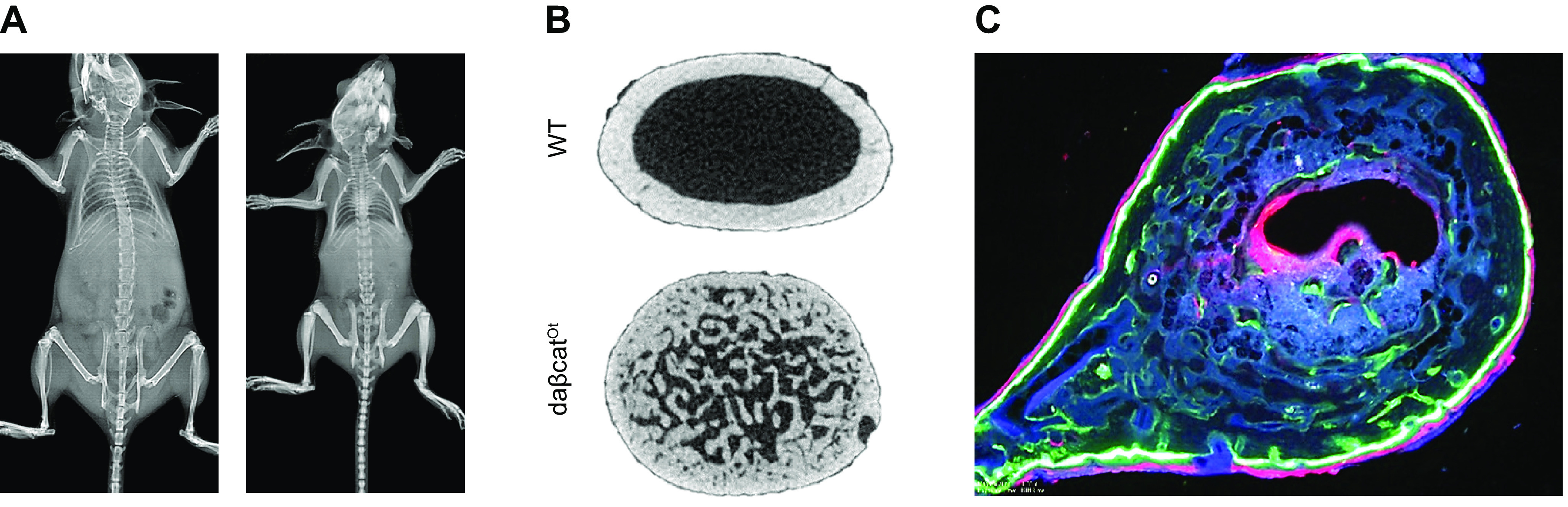
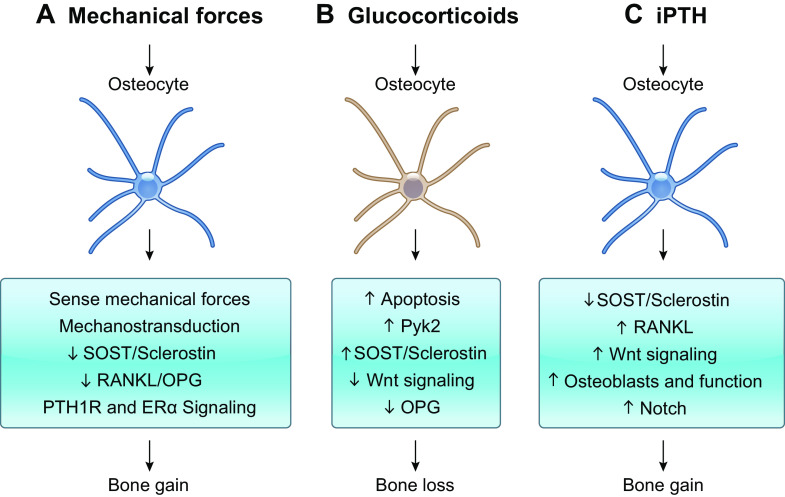
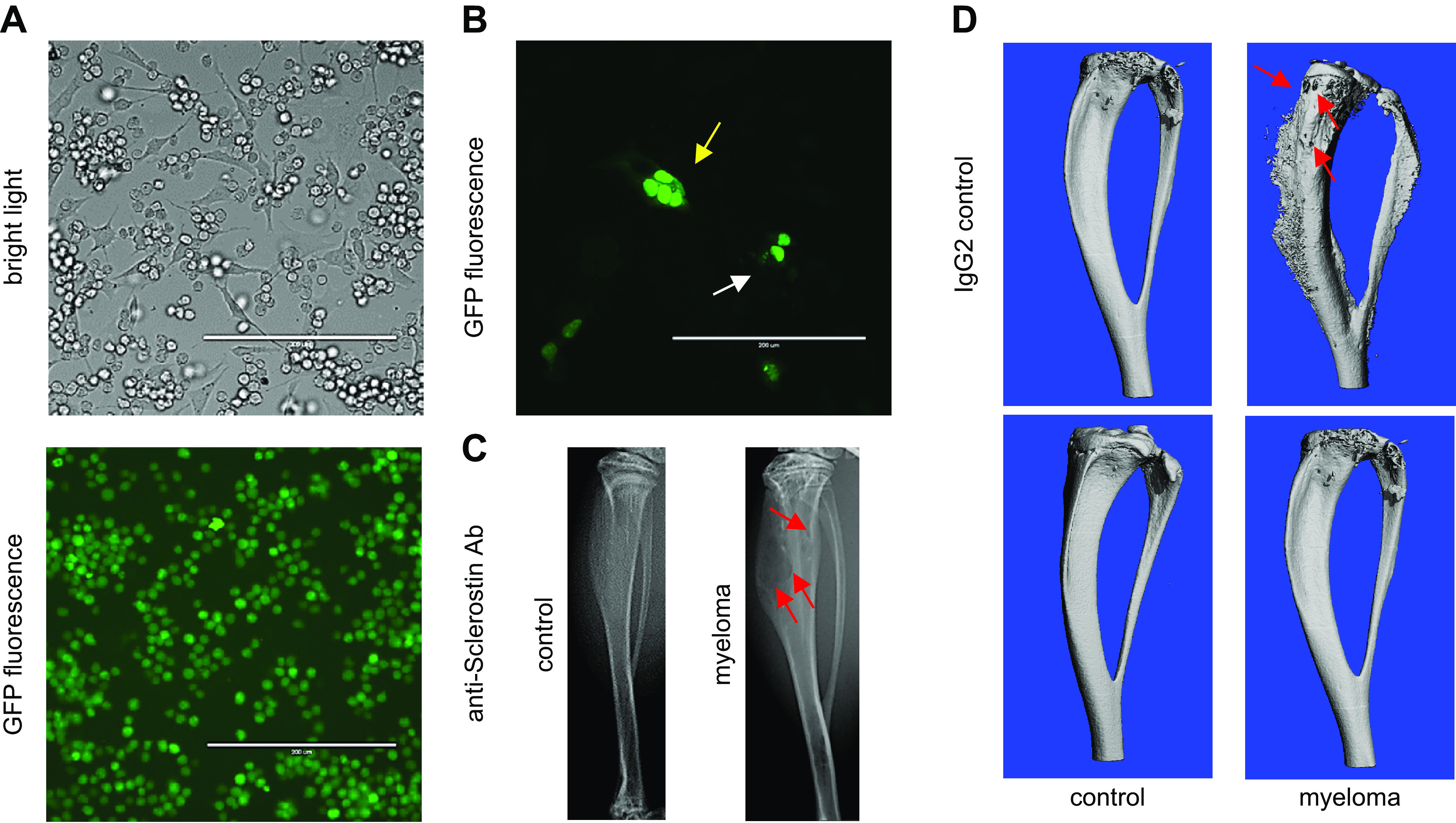
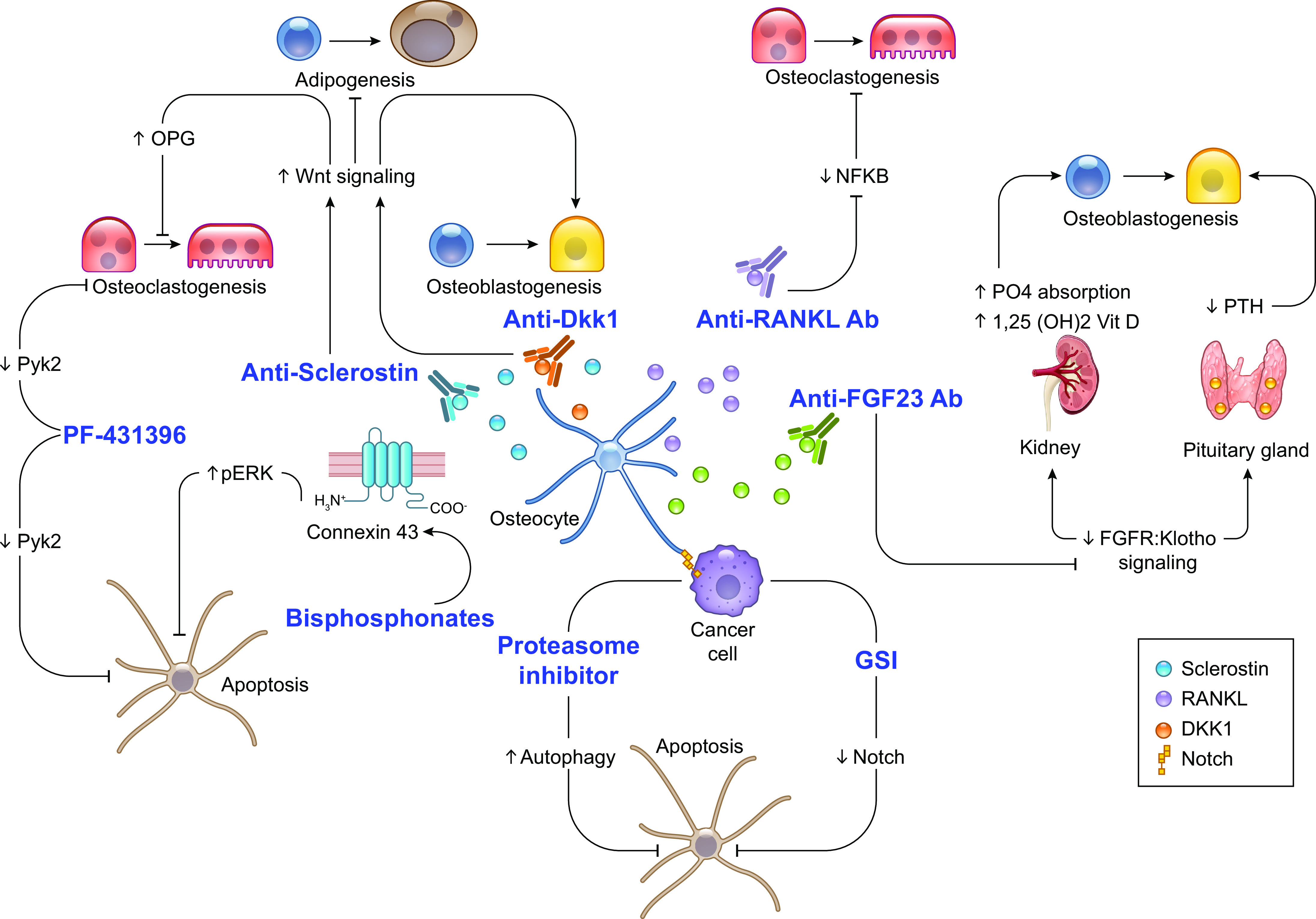
Osteocytes, former osteoblasts encapsulated by mineralized bone matrix, are far from being passive and metabolically inactive bone cells. Instead, osteocytes are multifunctional and dynamic cells capable of integrating hormonal and mechanical signals and transmitting them to effector cells in bone and in distant tissues. Osteocytes are a major source of molecules that regulate bone homeostasis by integrating both mechanical cues and hormonal signals that coordinate the differentiation and function of osteoclasts and osteoblasts. Osteocyte function is altered in both rare and common bone diseases, suggesting that osteocyte dysfunction is directly involved in the pathophysiology of several disorders affecting the skeleton. Advances in osteocyte biology initiated the development of novel therapeutics interfering with osteocyte-secreted molecules. Moreover, osteocytes are targets and key distributors of biological signals mediating the beneficial effects of several bone therapeutics used in the clinic. Here we review the most recent discoveries in osteocyte biology demonstrating that osteocytes regulate bone homeostasis and bone marrow fat via paracrine signaling, influence body composition and energy metabolism via endocrine signaling, and contribute to the damaging effects of diabetes mellitus and hematologic and metastatic cancers in the skeleton.
Keywords: Sclerostin; bone; cancer; hormonal signals; mechanical signals.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures



References
-
- Parfitt AM. Life history of osteocytes: relationship to bone age, bone remodeling, and bone fragility. J Musculoskelet Neuronal Interact 2: 499–500, 2002. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources