

A general approach to explore prokaryotic protein glycosylation reveals the unique surface layer modulation of an anammox bacterium
- PMID: 34341504
- PMCID: PMC8776859
- DOI: 10.1038/s41396-021-01073-y
A general approach to explore prokaryotic protein glycosylation reveals the unique surface layer modulation of an anammox bacterium
Abstract
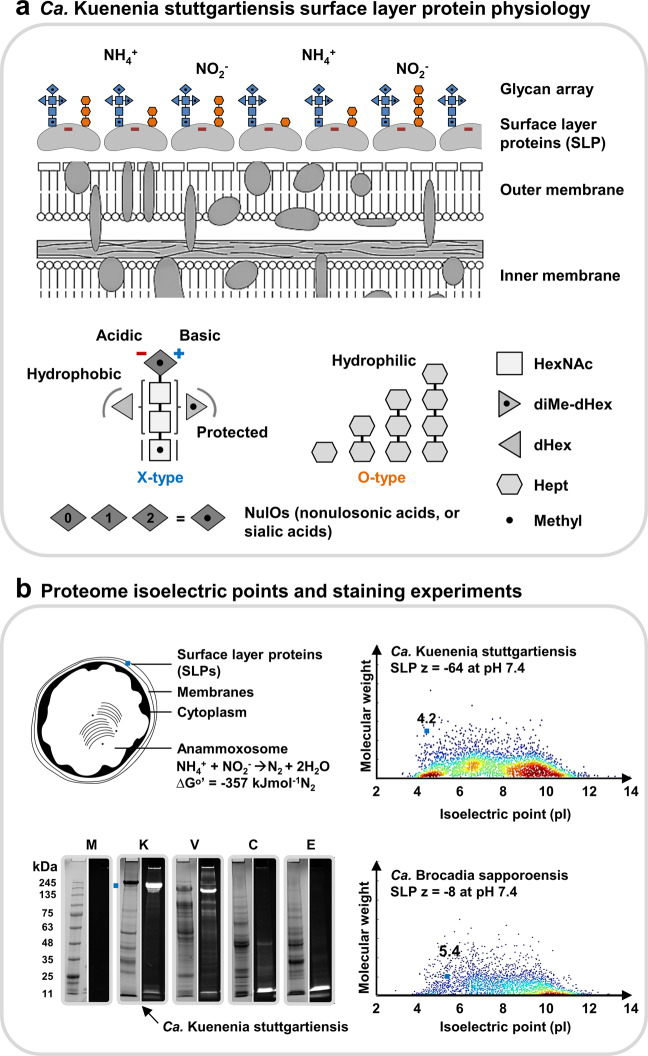
The enormous chemical diversity and strain variability of prokaryotic protein glycosylation makes their large-scale exploration exceptionally challenging. Therefore, despite the universal relevance of protein glycosylation across all domains of life, the understanding of their biological significance and the evolutionary forces shaping oligosaccharide structures remains highly limited. Here, we report on a newly established mass binning glycoproteomics approach that establishes the chemical identity of the carbohydrate components and performs untargeted exploration of prokaryotic oligosaccharides from large-scale proteomics data directly. We demonstrate our approach by exploring an enrichment culture of the globally relevant anaerobic ammonium-oxidizing bacterium Ca. Kuenenia stuttgartiensis. By doing so we resolve a remarkable array of oligosaccharides, which are produced by two seemingly unrelated biosynthetic routes, and which modify the same surface-layer protein simultaneously. More intriguingly, the investigated strain also accomplished modulation of highly specialized sugars, supposedly in response to its energy metabolism-the anaerobic oxidation of ammonium-which depends on the acquisition of substrates of opposite charges. Ultimately, we provide a systematic approach for the compositional exploration of prokaryotic protein glycosylation, and reveal a remarkable example for the evolution of complex oligosaccharides in bacteria.
© 2021. The Author(s), under exclusive licence to International Society for Microbial Ecology.
Conflict of interest statement
The authors declare no competing interests.
Figures
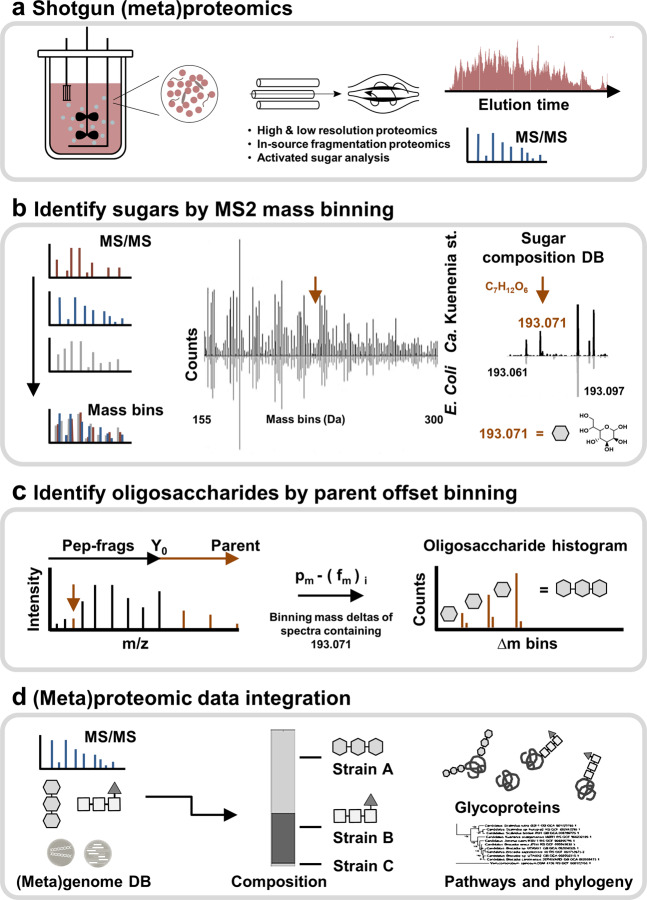
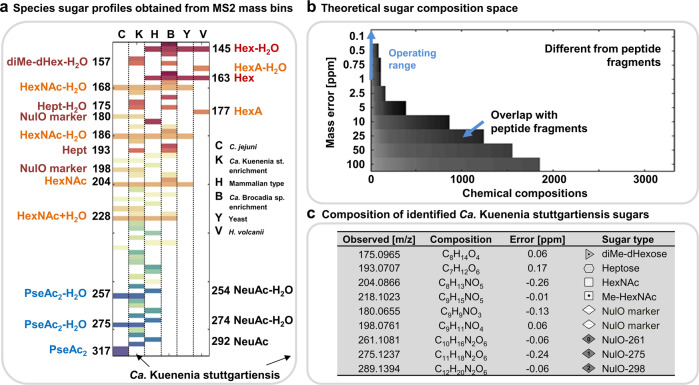
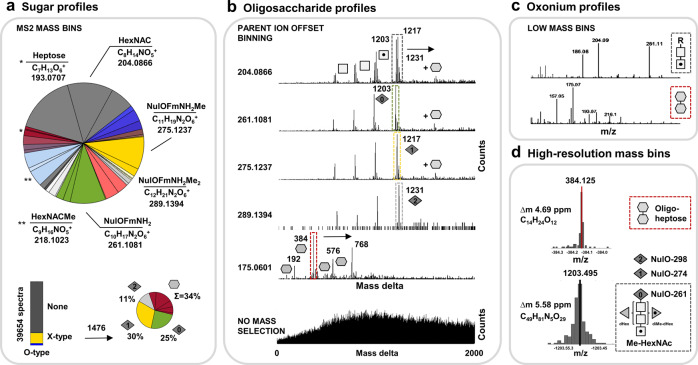
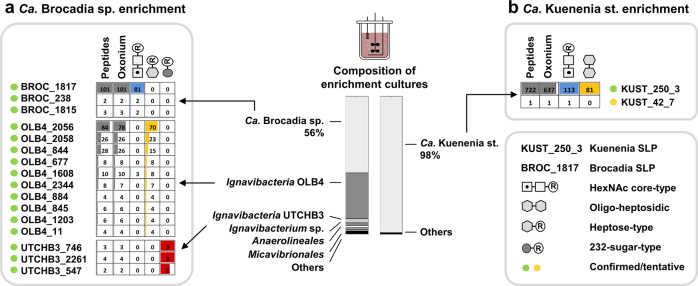
References
-
- den Ridder M, Daran-Lapujade P, Pabst M. Shot-gun proteomics: why thousands of unidentified signals matter. FEMS Yeast Res. 2020;20:foz088. - PubMed
-
- Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, Bertozzi CR, et al. Essentials of glycobiology. 3rd edition. (Cold Spring Harbor Laboratory Press, New York, 2015–2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
