

Vibrio cholerae Motility in Aquatic and Mucus-Mimicking Environments
- PMID: 34347522
- PMCID: PMC8478464
- DOI: 10.1128/AEM.01293-21
Vibrio cholerae Motility in Aquatic and Mucus-Mimicking Environments
Abstract
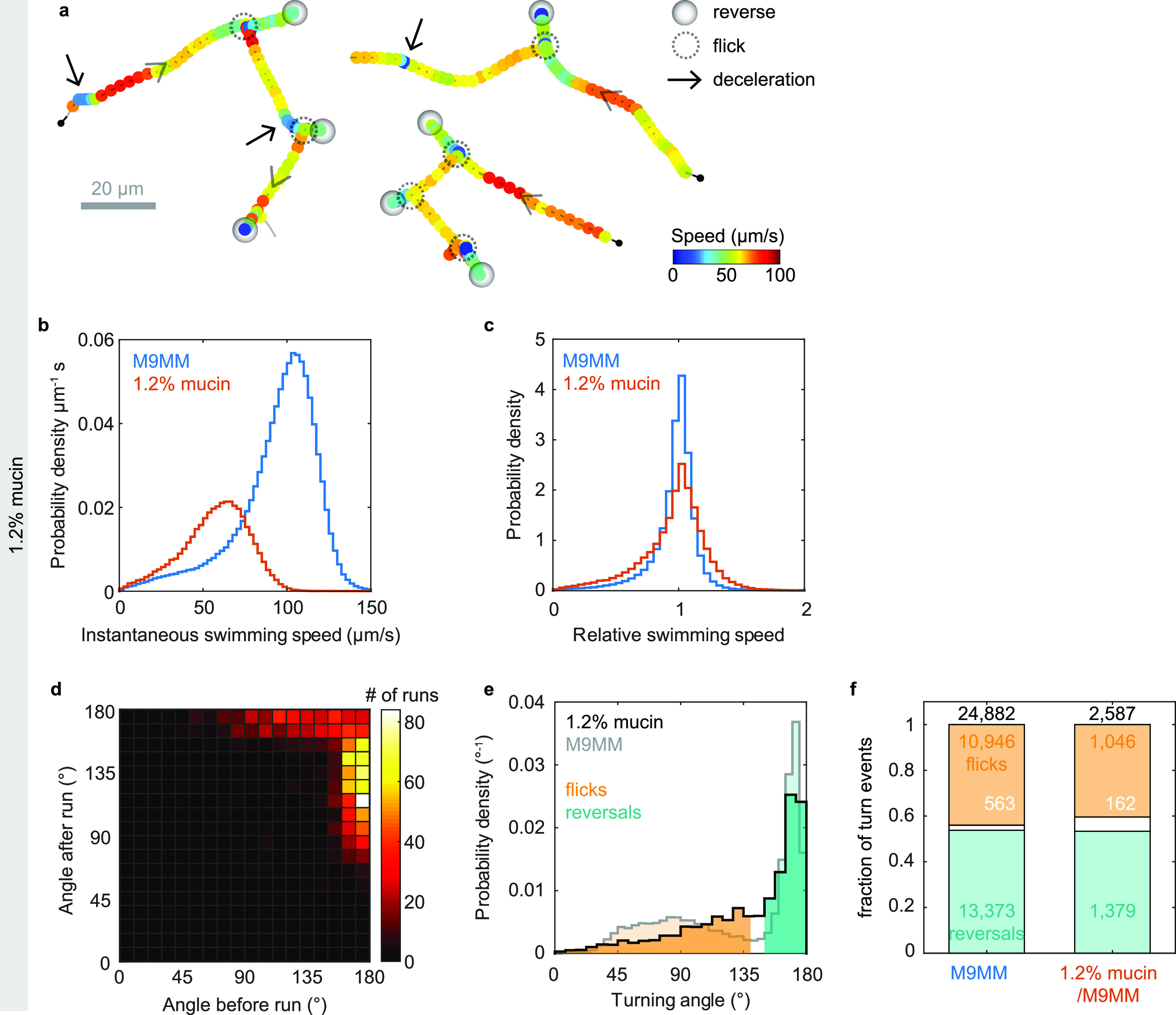
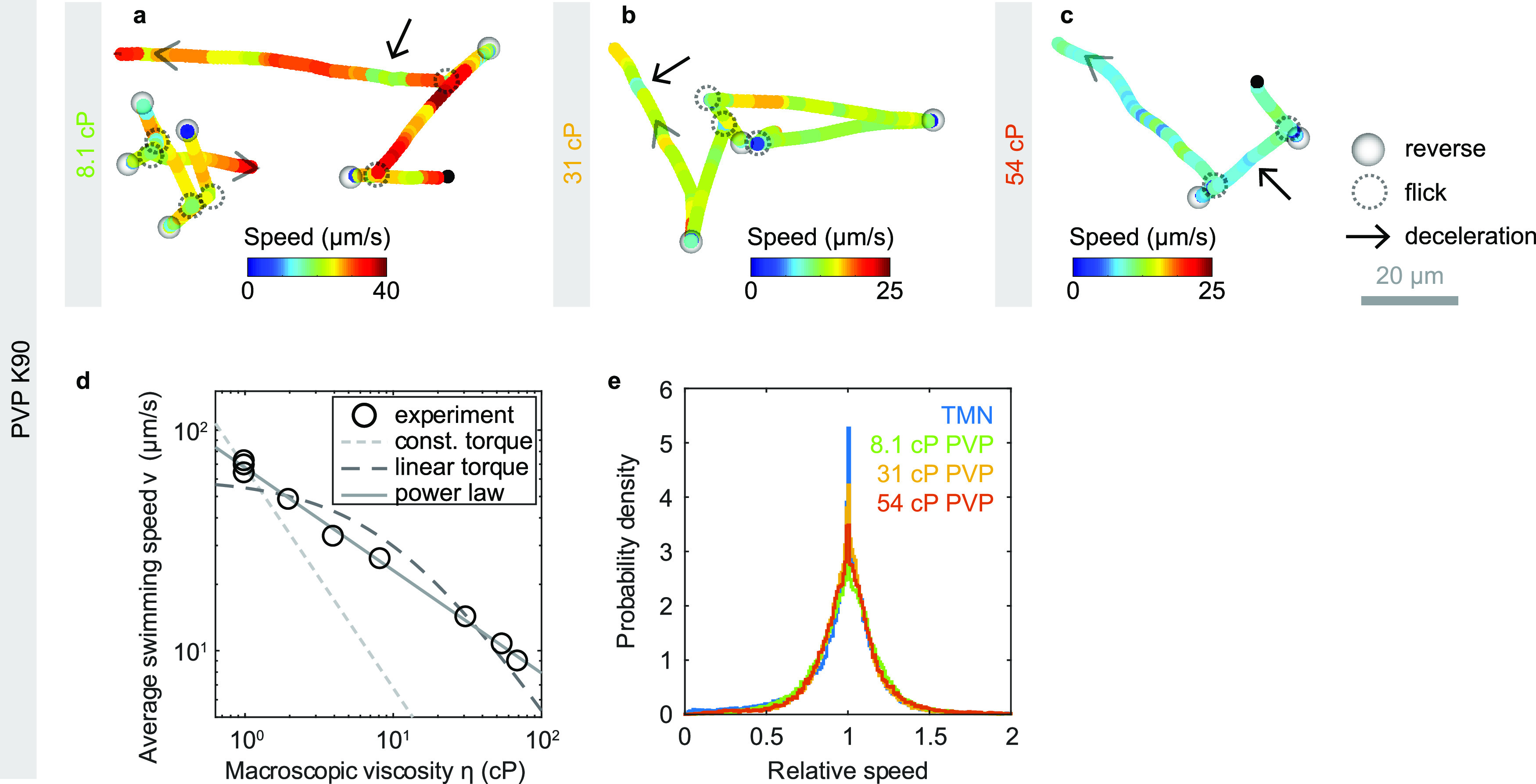
Cholera disease is caused by Vibrio cholerae infecting the lining of the small intestine and results in severe diarrhea. V. cholerae's swimming motility is known to play a crucial role in pathogenicity and may aid the bacteria in crossing the intestinal mucus barrier to reach sites of infection, but the exact mechanisms are unknown. The cell can be either pushed or pulled by its single polar flagellum, but there is no consensus on the resulting repertoire of motility behaviors. We use high-throughput three-dimensional (3D) bacterial tracking to observe V. cholerae swimming in buffer, in viscous solutions of the synthetic polymer PVP, and in mucin solutions that may mimic the host environment. We perform a statistical characterization of its motility behavior on the basis of large 3D trajectory data sets. We find that V. cholerae performs asymmetric run-reverse-flick motility, consisting of a sequence of a forward run, reversal, and a shorter backward run, followed by a turn by approximately 90°, called a flick, preceding the next forward run. Unlike many run-reverse-flick swimmers, V. cholerae's backward runs are much shorter than its forward runs, resulting in an increased effective diffusivity. We also find that the swimming speed is not constant but subject to frequent decreases. The turning frequency in mucin matches that observed in buffer. Run-reverse-flick motility and speed fluctuations are present in all environments studied, suggesting that these behaviors also occur in natural aquatic habitats as well as the host environment. IMPORTANCE Cholera disease produces vomiting and severe diarrhea and causes approximately 100,000 deaths per year worldwide. The disease is caused by the bacterium Vibrio cholerae colonizing the lining of the small intestine. V. cholerae's ability to swim is known to increase its infectivity, but the underlying mechanisms are not known. One possibility is that swimming aids in crossing the protective mucus barrier that covers the lining of the small intestine. Our work characterizing how V. cholerae swims in environments that mimic properties of the host environment may advance the understanding of how motility contributes to infection.
Keywords: Vibrio cholerae; flagellar motility; mucin.
Figures
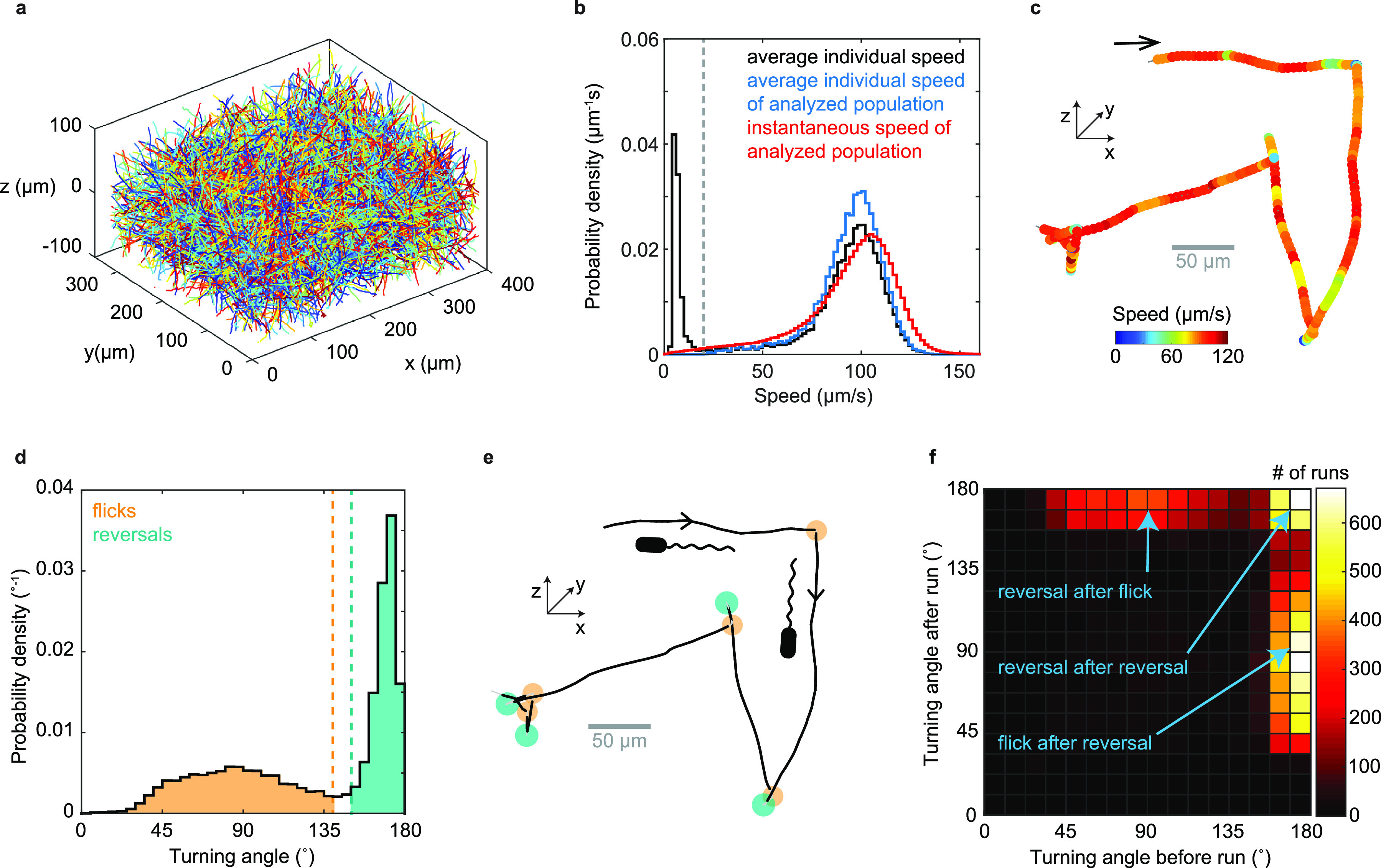
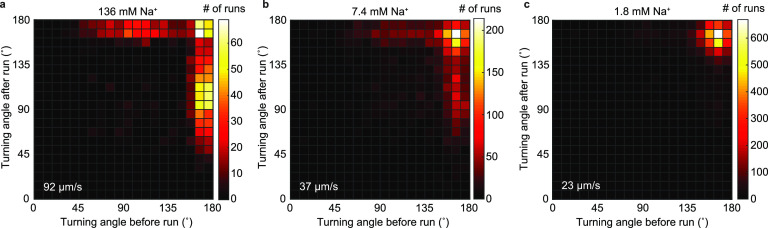
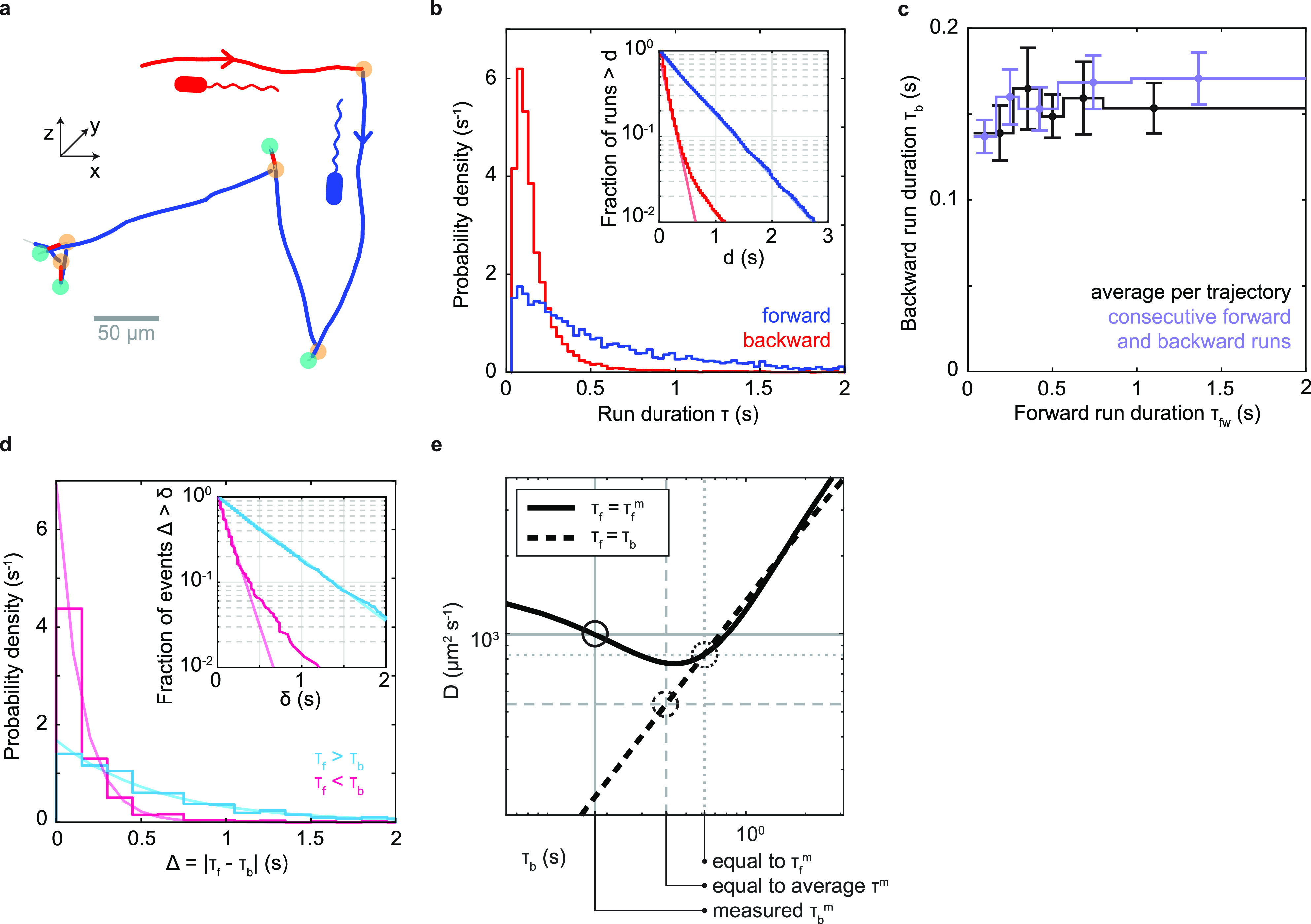
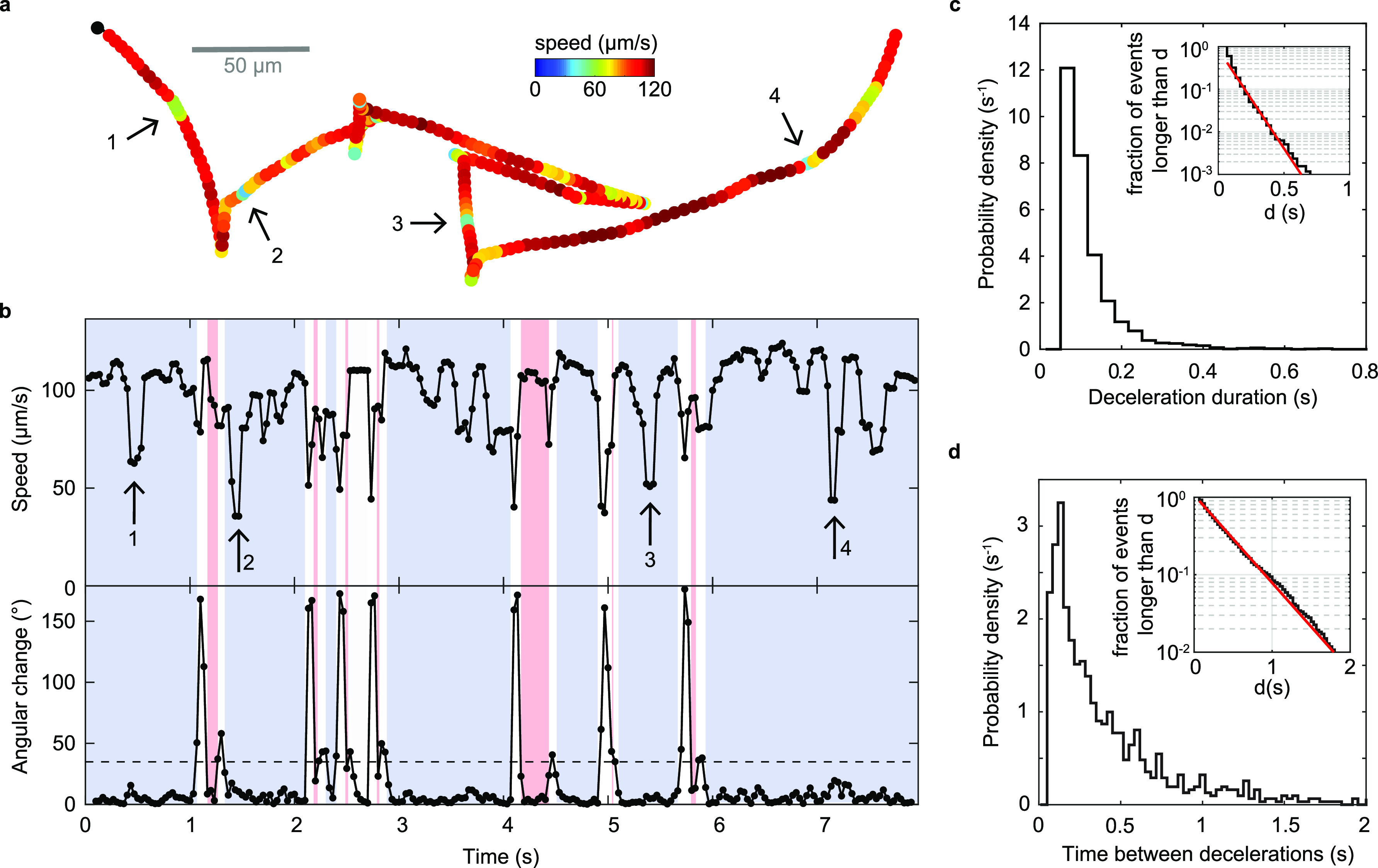
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
