

Ketogenesis impact on liver metabolism revealed by proteomics of lysine β-hydroxybutyrylation
- PMID: 34348140
- PMCID: PMC8372761
- DOI: 10.1016/j.celrep.2021.109487
Ketogenesis impact on liver metabolism revealed by proteomics of lysine β-hydroxybutyrylation
Abstract
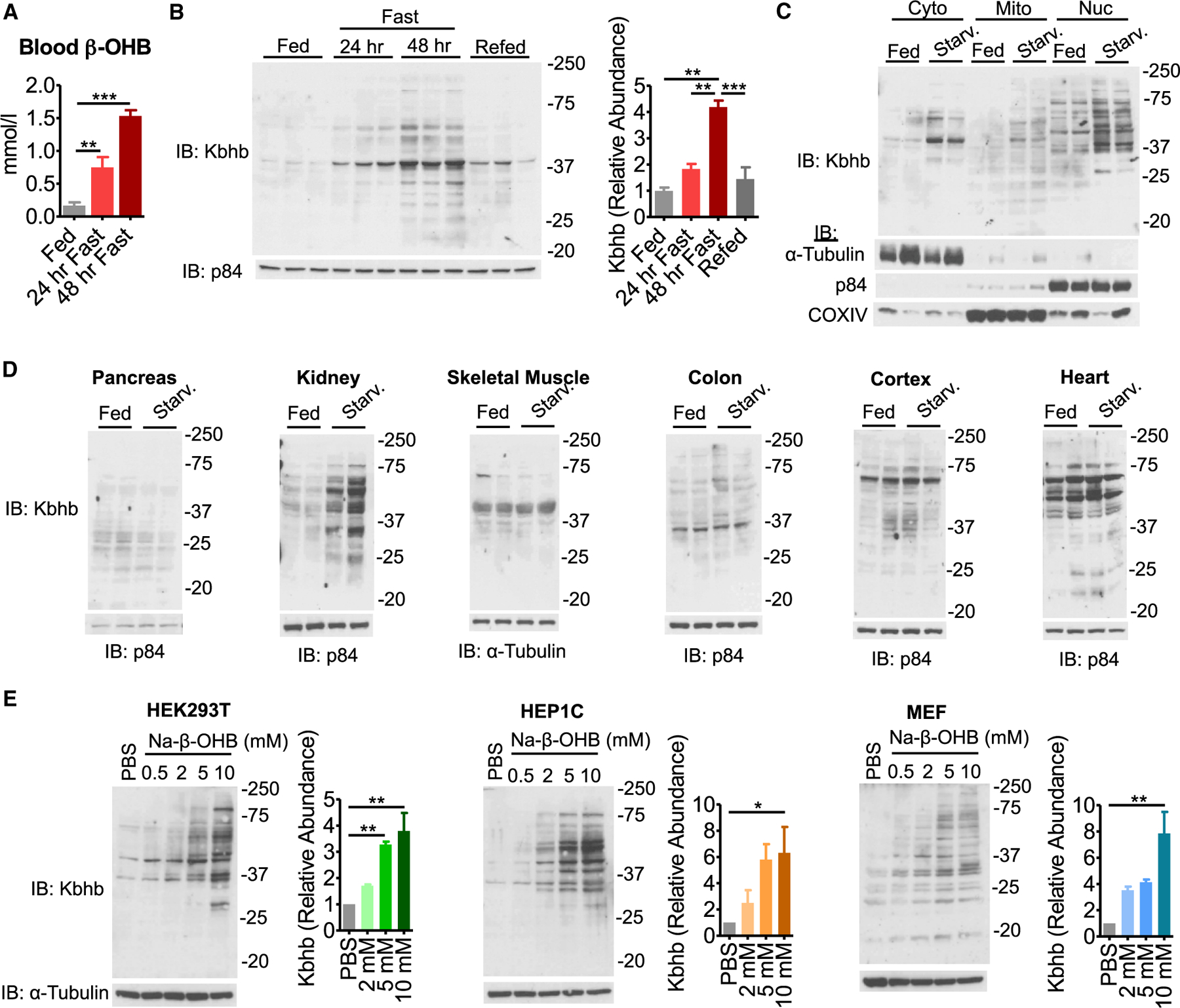
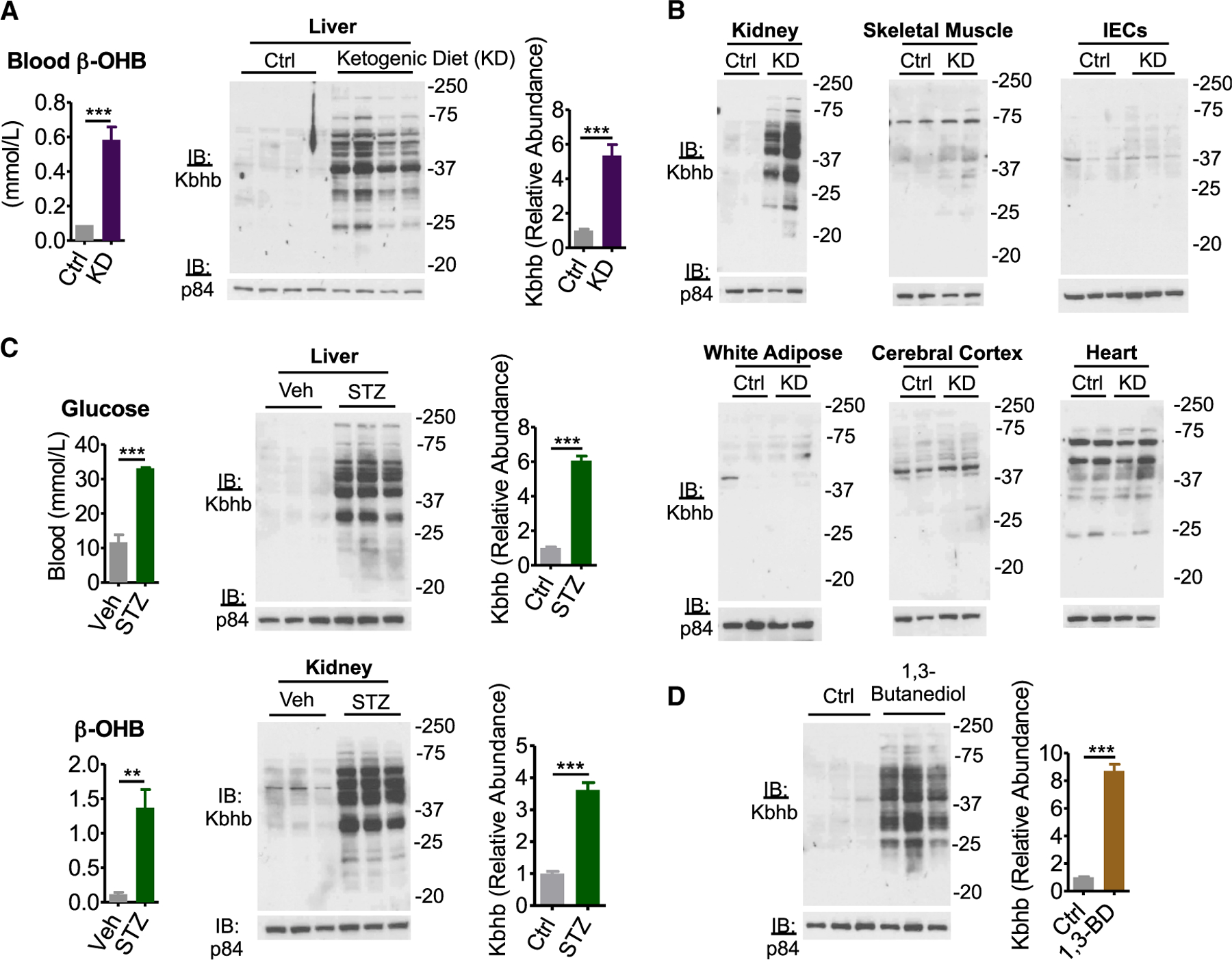
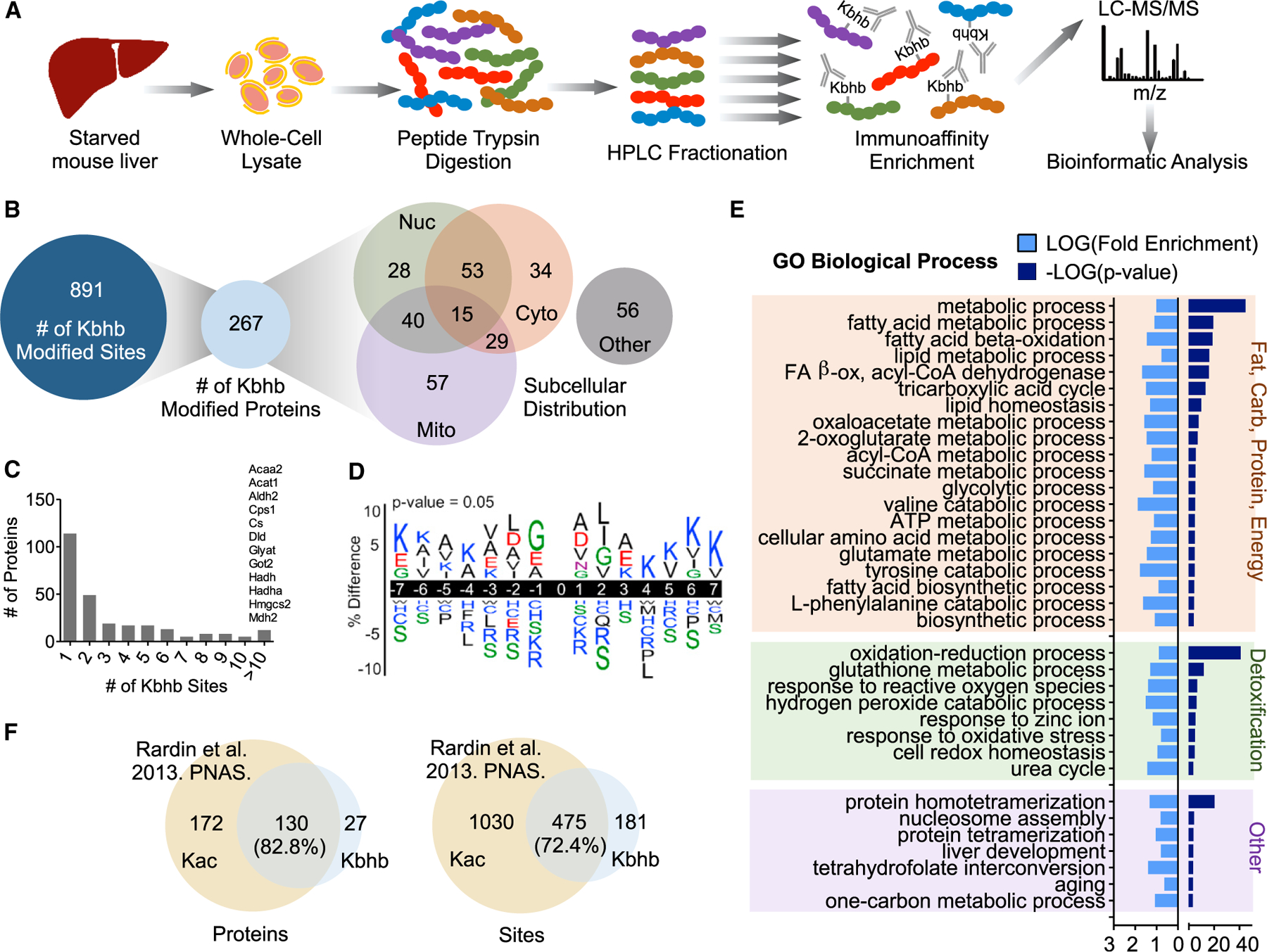
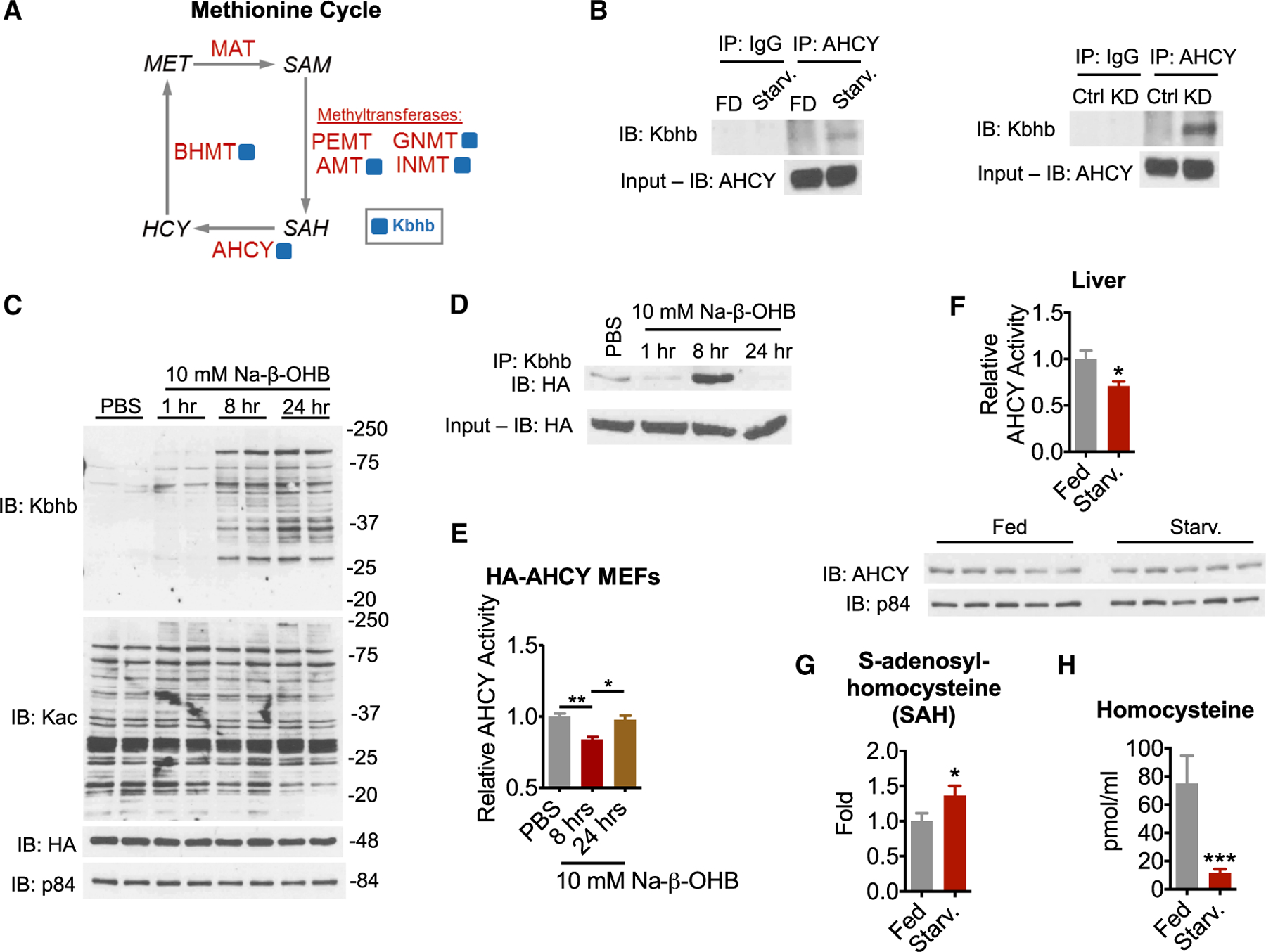
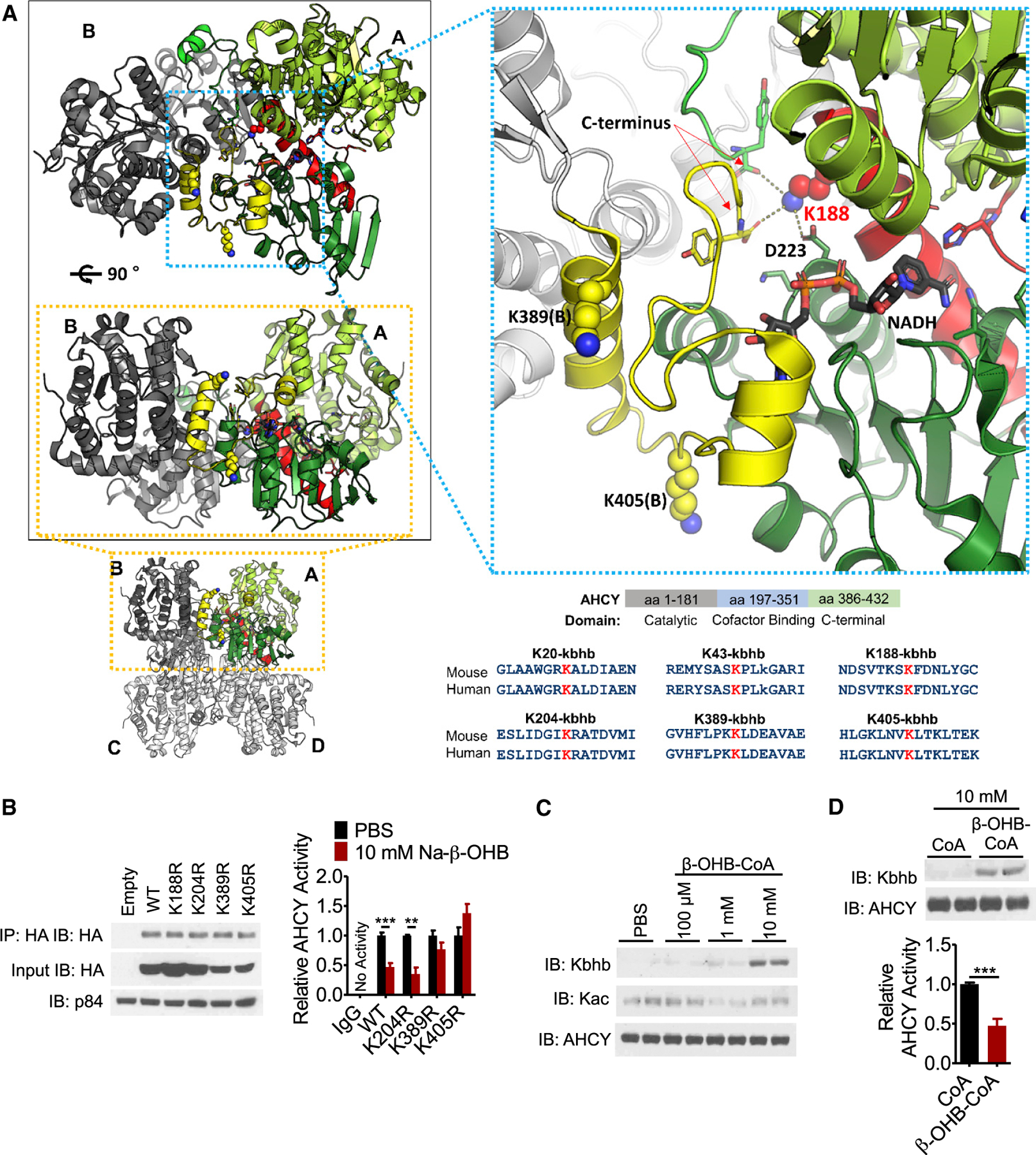
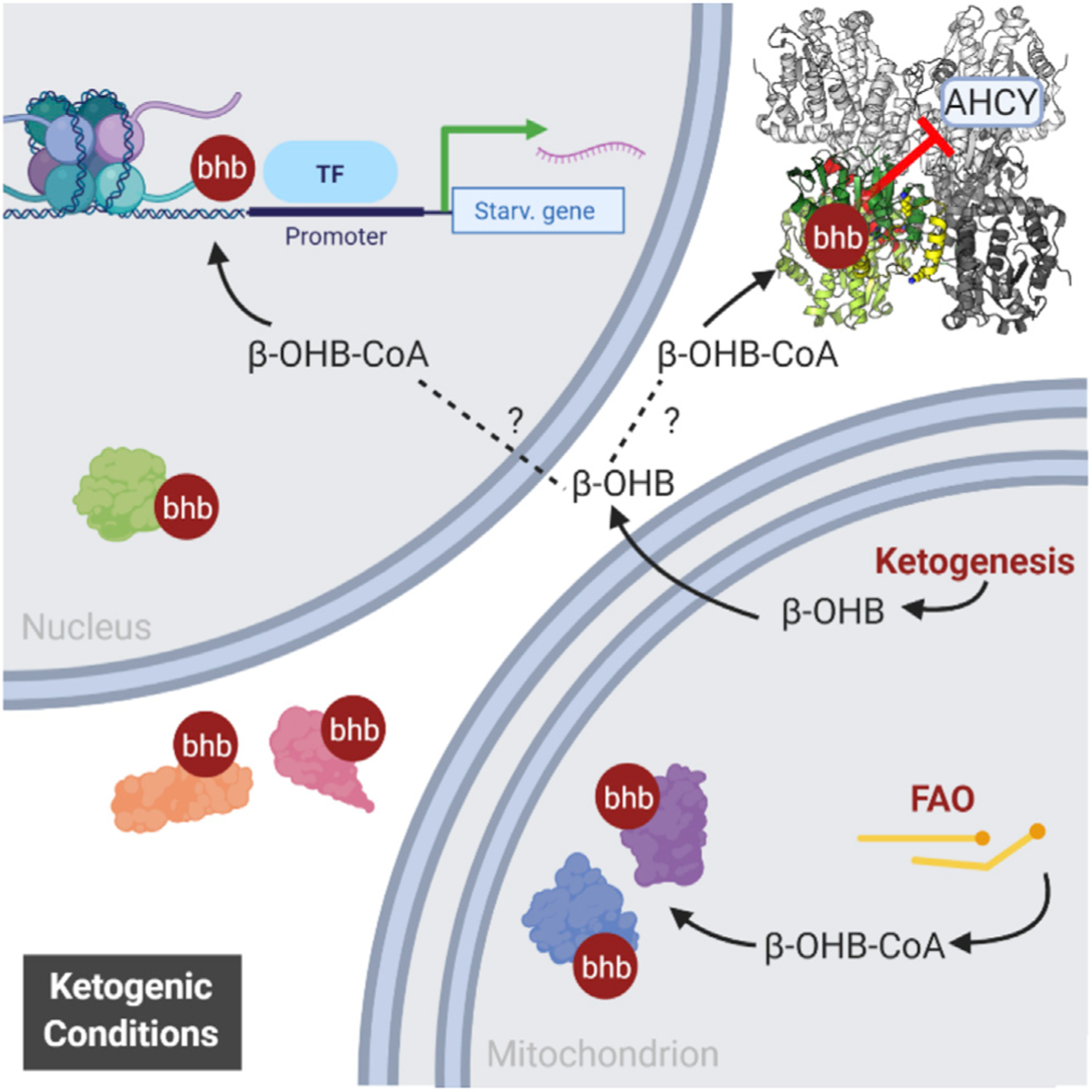
Ketone bodies are bioactive metabolites that function as energy substrates, signaling molecules, and regulators of histone modifications. β-hydroxybutyrate (β-OHB) is utilized in lysine β-hydroxybutyrylation (Kbhb) of histones, and associates with starvation-responsive genes, effectively coupling ketogenic metabolism with gene expression. The emerging diversity of the lysine acylation landscape prompted us to investigate the full proteomic impact of Kbhb. Global protein Kbhb is induced in a tissue-specific manner by a variety of interventions that evoke β-OHB. Mass spectrometry analysis of the β-hydroxybutyrylome in mouse liver revealed 891 sites of Kbhb within 267 proteins enriched for fatty acid, amino acid, detoxification, and one-carbon metabolic pathways. Kbhb inhibits S-adenosyl-L-homocysteine hydrolase (AHCY), a rate-limiting enzyme of the methionine cycle, in parallel with altered metabolite levels. Our results illuminate the role of Kbhb in hepatic metabolism under ketogenic conditions and demonstrate a functional consequence of this modification on a central metabolic enzyme.
Keywords: AHCY; S-adenosyl-L-homocysteine hydrolase; ketogenesis; ketogenic diet; liver metabolism; lysine acylation; methionine cycle; β-hydroxybutyrate; β-hydroxybutyrylation.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Ketogenic modification of the liver proteome.Nat Rev Endocrinol. 2021 Nov;17(11):638. doi: 10.1038/s41574-021-00560-8. Nat Rev Endocrinol. 2021. PMID: 34446929 No abstract available.
References
-
- Balasse EO, and Féry F (1989). Ketone body production and disposal: effects of fasting, diabetes, and exercise. Diabetes Metab. Rev 5, 247–270. - PubMed
-
- Barić I (2009). Inherited disorders in the conversion of methionine to homocysteine. J. Inherit. Metab. Dis 32, 459–471. - PubMed
-
- Cahill GF Jr. (2006). Fuel metabolism in starvation. Annu. Rev. Nutr 26, 1–22. - PubMed
