

A widely diverged locus involved in locomotor adaptation in Heliconius butterflies
- PMID: 34348900
- PMCID: PMC8336958
- DOI: 10.1126/sciadv.abh2340
A widely diverged locus involved in locomotor adaptation in Heliconius butterflies
Abstract
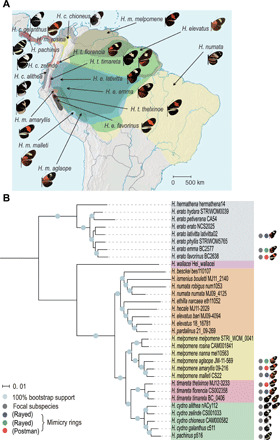
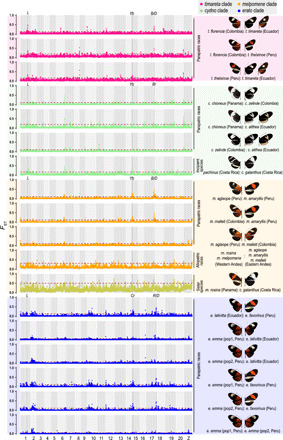
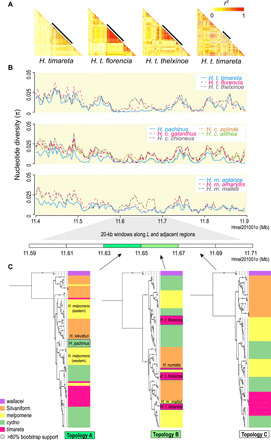
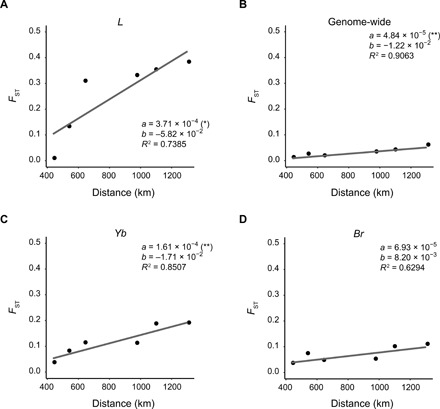
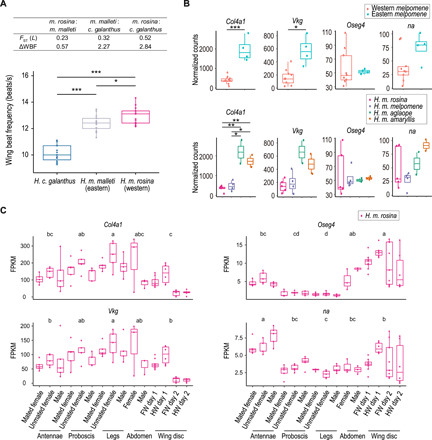
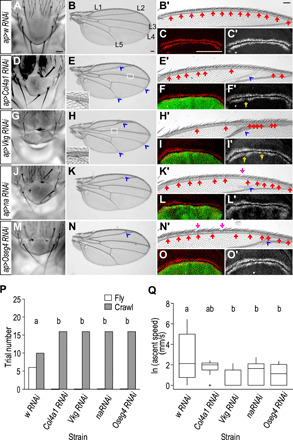
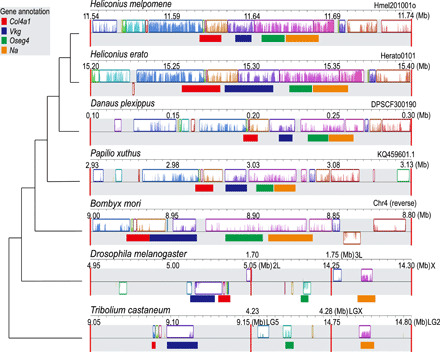
Heliconius butterflies have undergone adaptive radiation and therefore serve as an excellent system for exploring the continuum of speciation and adaptive evolution. However, there is a long-lasting paradox between their convergent mimetic wing patterns and rapid divergence in speciation. Here, we characterize a locus that consistently displays high divergence among Heliconius butterflies and acts as an introgression hotspot. We further show that this locus contains multiple genes related to locomotion and conserved in Lepidoptera. In light of these findings, we consider that locomotion traits may be under selection, and if these are heritable traits that are selected for, then they might act as species barriers.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Lamichhaney S., Berglund J., Almén M. S., Maqbool K., Grabherr M., Martinez-Barrio A., Promerová M., Rubin C. J., Wang C., Zamani N., Grant B. R., Grant P. R., Webster M. T., Andersson L., Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature 518, 371–375 (2015). - PubMed
-
- Peichel C. L., Nereng K. S., Ohgi K. A., Cole B. L. E., Colosimo P. F., Buerkle C. A., Schluter D., Kingsley D. M., The genetic architecture of divergence between threespine stickleback species. Nature 414, 901–905 (2001). - PubMed
-
- Servedio M. R., Van Doorn G. S., Kopp M., Frame A. M., Nosil P., Magic traits in speciation: ‘Magic’ but not rare? Trends Ecol. Evol. 26, 389–397 (2011). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
