

Deciphering the biology of KIR2DL3+ T lymphocytes that are associated to relapse in haploidentical HSCT
- PMID: 34349169
- PMCID: PMC8338934
- DOI: 10.1038/s41598-021-95245-7
Deciphering the biology of KIR2DL3+ T lymphocytes that are associated to relapse in haploidentical HSCT
Abstract
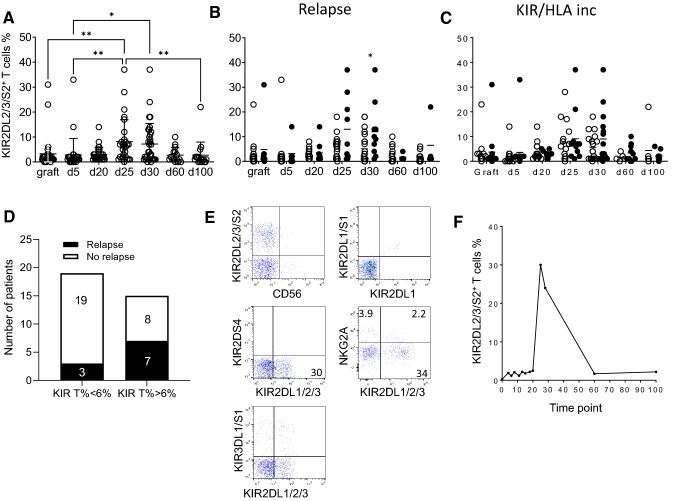
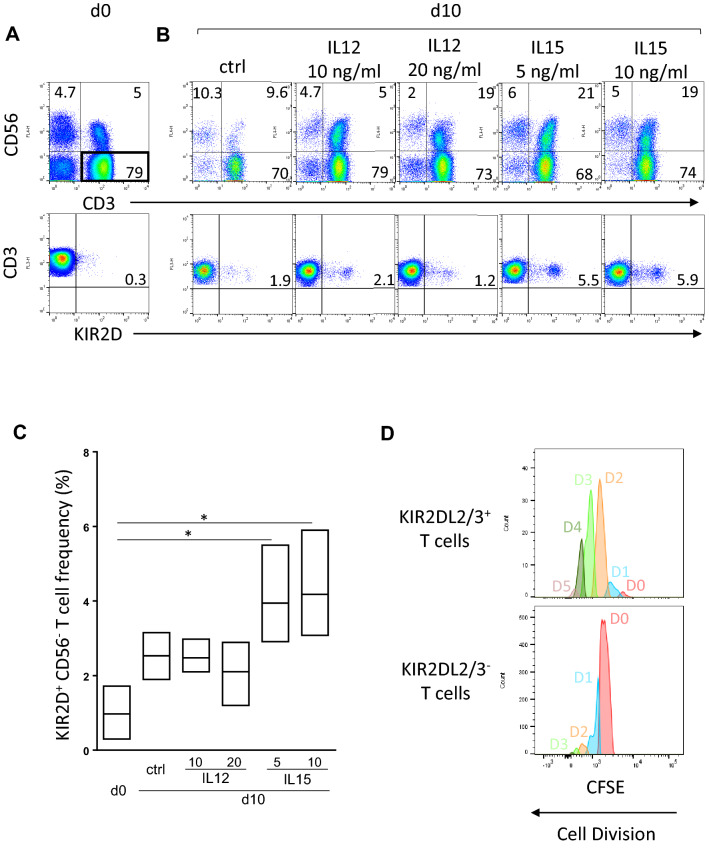
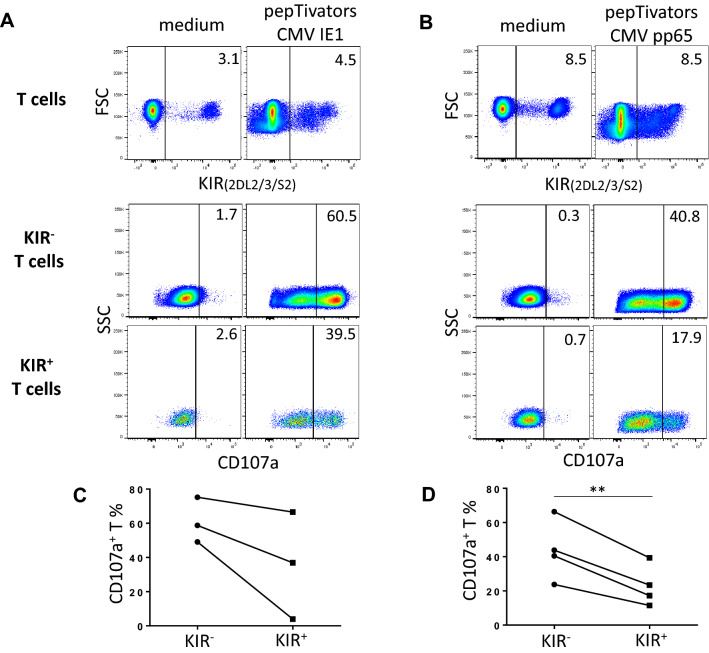
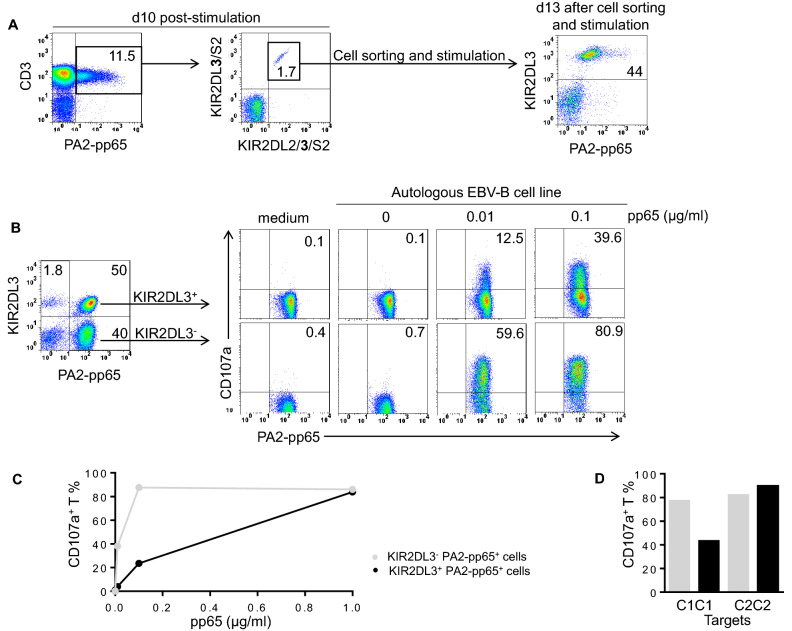
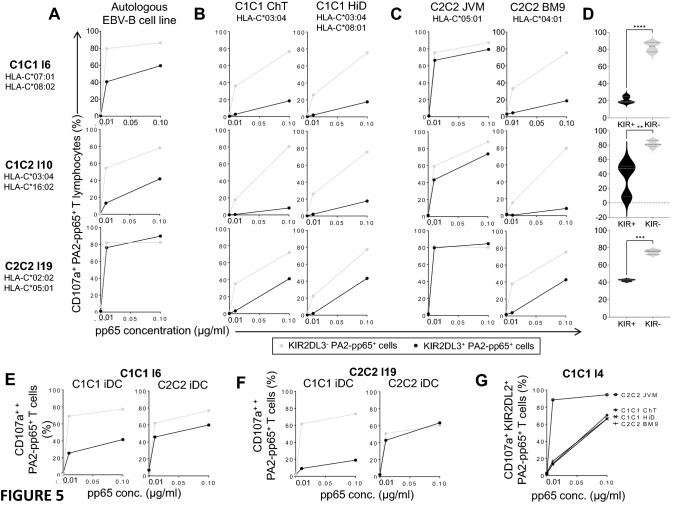
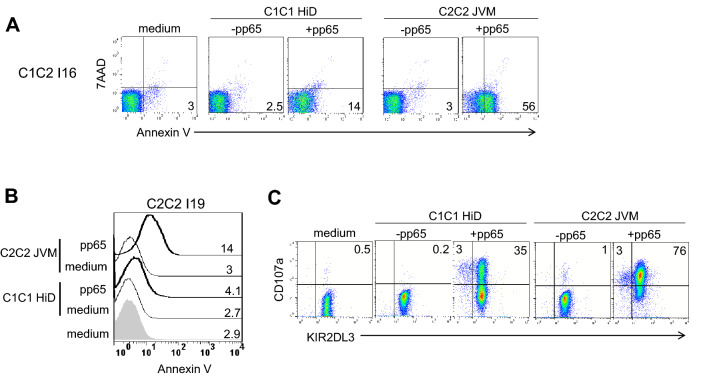
KIR are mainly expressed on NK cells and to a lesser extent on T lymphocytes. Although the KIR NK cell repertoire was well explored in haploidentical Hematopoietic Stem Cell Transplantation (HSCT), KIR T cell compartment remains to be investigated in this context. In this study, the investigation of NK receptors on T lymphocytes during immune reconstitution after T-cell-replete haploidentical HSCT with Post-Transplant Cyclophosphamide (PTCy) has shown a significant increase of KIR2DL2/3+ T cell frequency at day 25. This was especially observed at day 30 in recipients who relapsed. IL-15 but not IL-12 increased in vitro KIR+ T cell expansion suggesting that the raised IL-15 serum concentration observed after PTCy in haploidentical HSCT might increase KIR+ T cell frequency. Moreover, investigations from healthy blood donors showed a higher inhibiting effect of KIR2DL3 on CMV specific T cell response against allogeneic than autologous C1+ target cells. The association of KIR+ T cell subset with relapse may suggest that inhibitory KIR2DL2/3 limit anti-leukemic effect of specific T lymphocytes at this early step of immune reconstitution. Further phenotypic and mechanistic investigations on this cell subset from a broader cohort of HSCT recipients should clarify its potential implication in relapse occurrence. Our results demonstrate that KIR-HLA interactions known to modulate NK cell functions also modulate T cell immune responses in the context of allogeneic HSCT.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Mandelboim O, et al. Protection from lysis by natural killer cells of group 1 and 2 specificity is mediated by residue 80 in human histocompatibility leukocyte antigen C alleles and also occurs with empty major histocompatibility complex molecules. J. Exp. Med. 1996;184:913–922. doi: 10.1084/jem.184.3.913. - DOI - PMC - PubMed
-
- Pende D, et al. Anti-leukemia activity of alloreactive NK cells in KIR ligand-mismatched haploidentical HSCT for pediatric patients: Evaluation of the functional role of activating KIR and redefinition of inhibitory KIR specificity. Blood. 2009;113:3119–3129. doi: 10.1182/blood-2008-06-164103. - DOI - PubMed
-
- David G, et al. Large spectrum of HLA-C recognition by killer Ig-like receptor (KIR)2DL2 and KIR2DL3 and restricted C1 SPECIFICITY of KIR2DS2: Dominant impact of KIR2DL2/KIR2DS2 on KIR2D NK cell repertoire formation. J. Immunol. Baltim. 2013;191:4778–4788. doi: 10.4049/jimmunol.1301580. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
