

Activation of IRE1, PERK and salt-inducible kinases leads to Sec body formation in Drosophila S2 cells
- PMID: 34350957
- PMCID: PMC8445602
- DOI: 10.1242/jcs.258685
Activation of IRE1, PERK and salt-inducible kinases leads to Sec body formation in Drosophila S2 cells
Abstract
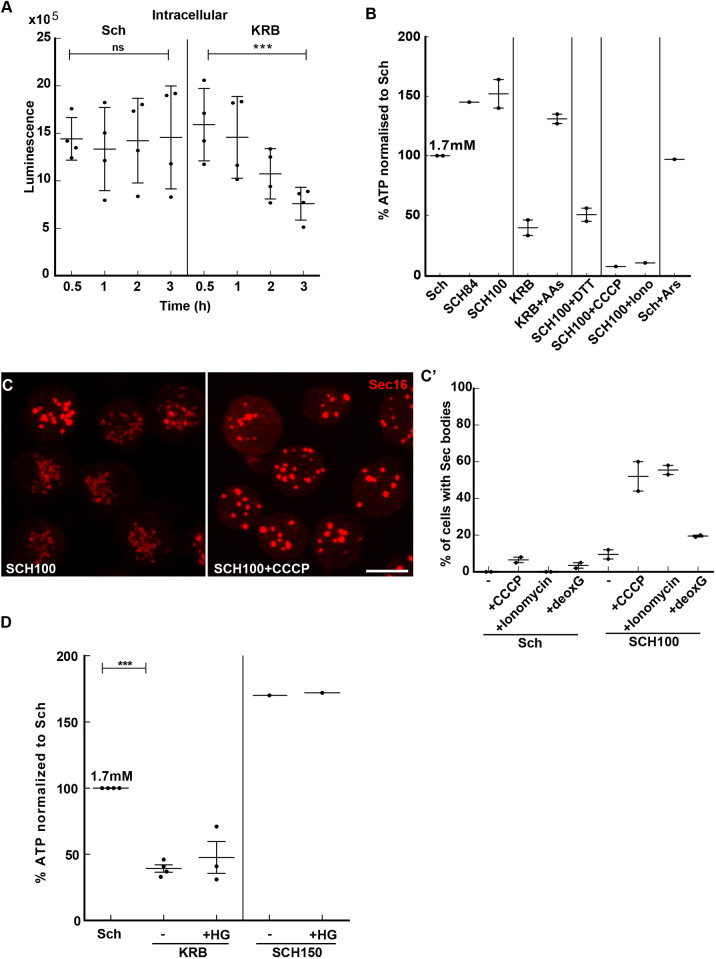
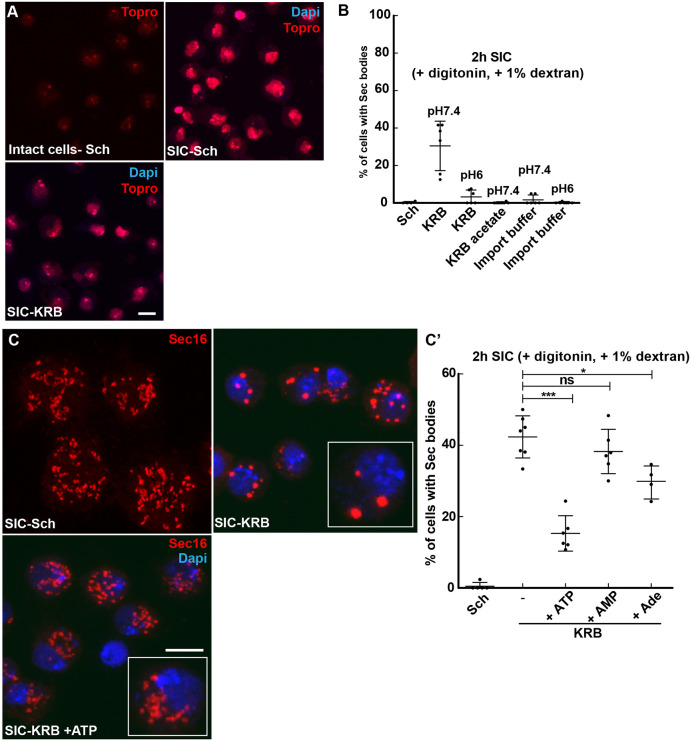
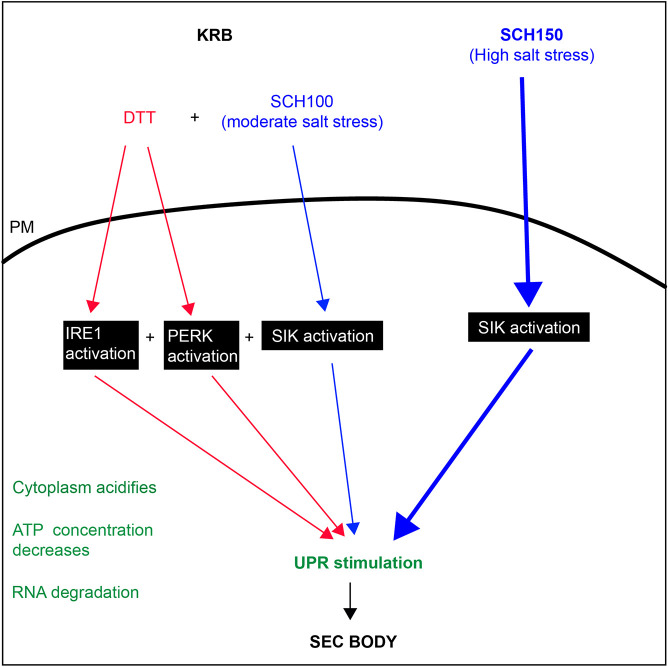
The phase separation of the non-membrane bound Sec bodies occurs in Drosophila S2 cells by coalescence of components of the endoplasmic reticulum (ER) exit sites under the stress of amino acid starvation. Here, we address which signaling pathways cause Sec body formation and find that two pathways are critical. The first is the activation of the salt-inducible kinases (SIKs; SIK2 and SIK3) by Na+ stress, which, when it is strong, is sufficient. The second is activation of IRE1 and PERK (also known as PEK in flies) downstream of ER stress induced by the absence of amino acids, which needs to be combined with moderate salt stress to induce Sec body formation. SIK, and IRE1 and PERK activation appear to potentiate each other through the stimulation of the unfolded protein response, a key parameter in Sec body formation. This work shows the role of SIKs in phase transition and re-enforces the role of IRE1 and PERK as a metabolic sensor for the level of circulating amino acids and salt. This article has an associated First Person interview with the first author of the paper.
Keywords: Drosophila S2 cells; Amino acid starvation; Phase separation; Salt stress; Sec body; Unfolded protein response.
© 2021. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
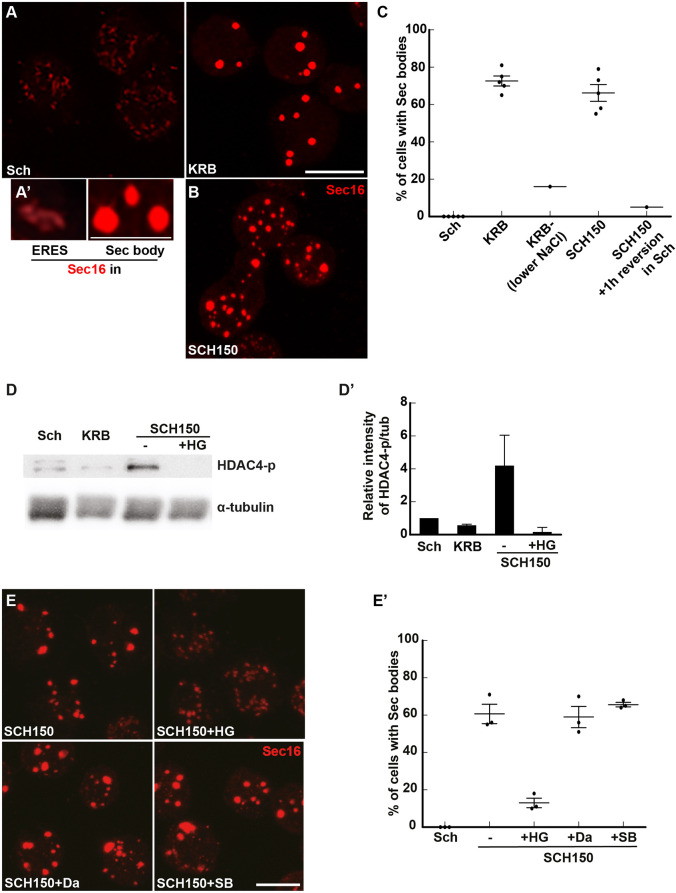
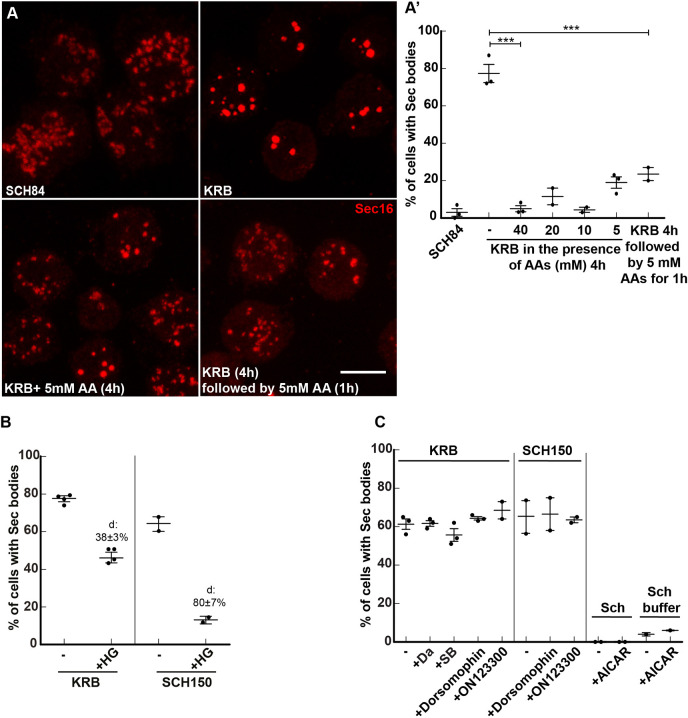
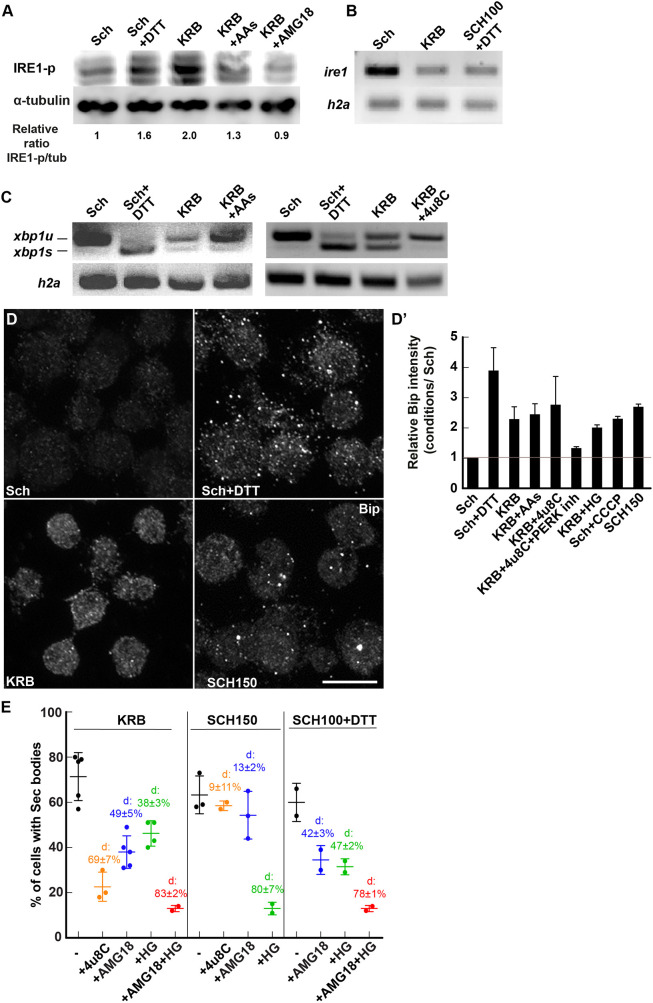
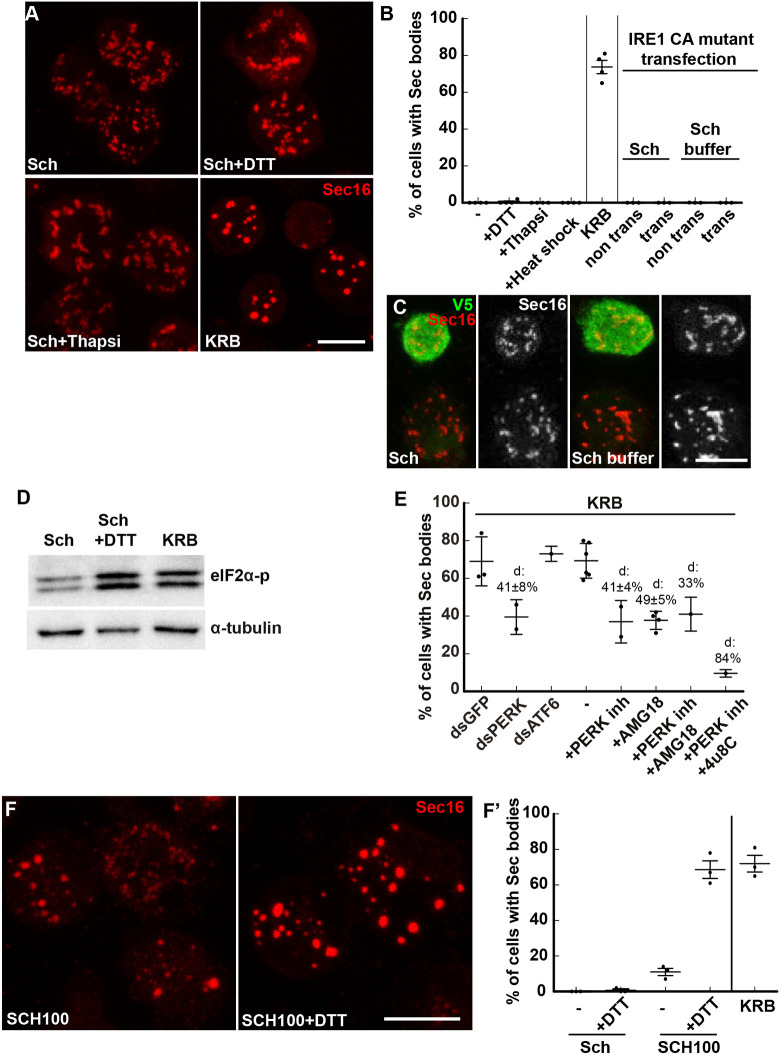
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
