

Butyrate and the Intestinal Epithelium: Modulation of Proliferation and Inflammation in Homeostasis and Disease
- PMID: 34359944
- PMCID: PMC8304699
- DOI: 10.3390/cells10071775
Butyrate and the Intestinal Epithelium: Modulation of Proliferation and Inflammation in Homeostasis and Disease
Abstract
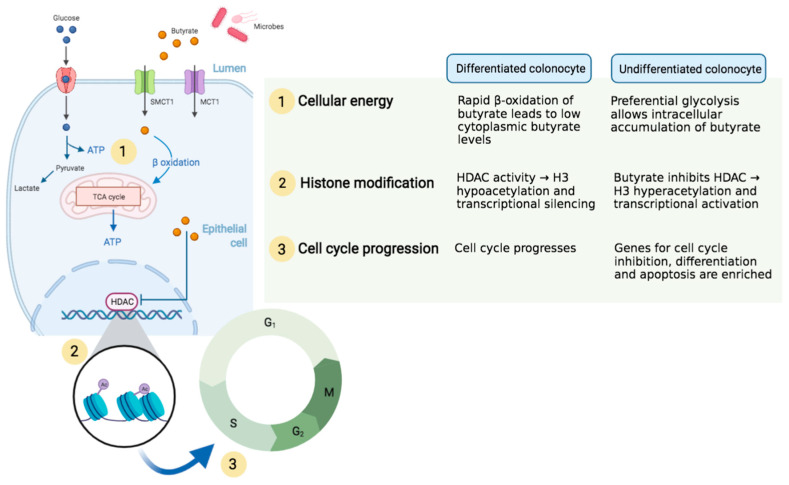
The microbial metabolite butyrate serves as a link between the intestinal microbiome and epithelium. The monocarboxylate transporters MCT1 and SMCT1 are the predominant means of butyrate transport from the intestinal lumen to epithelial cytoplasm, where the molecule undergoes rapid β-oxidation to generate cellular fuel. However, not all epithelial cells metabolize butyrate equally. Undifferentiated colonocytes, including neoplastic cells and intestinal stem cells at the epithelial crypt base preferentially utilize glucose over butyrate for cellular fuel. This divergent metabolic conditioning is central to the phenomenon known as "butyrate paradox", in which butyrate induces contradictory effects on epithelial proliferation in undifferentiated and differentiated colonocytes. There is evidence that accumulation of butyrate in epithelial cells results in histone modification and altered transcriptional activation that halts cell cycle progression. This manifests in the apparent protective effect of butyrate against colonic neoplasia. A corollary to this process is butyrate-induced inhibition of intestinal stem cells. Yet, emerging research has illustrated that the evolution of the crypt, along with butyrate-producing bacteria in the intestine, serve to protect crypt base stem cells from butyrate's anti-proliferative effects. Butyrate also regulates epithelial inflammation and tolerance to antigens, through production of anti-inflammatory cytokines and induction of tolerogenic dendritic cells. The role of butyrate in the pathogenesis and treatment of intestinal neoplasia, inflammatory bowel disease and malabsorptive states is evolving, and holds promise for the potential translation of butyrate's cellular function into clinical therapies.
Keywords: butyrate; intestinal epithelium; microbiome; short chain fatty acid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Louis P., Hold G.L., Flint H.J. The gut microbiota, bacterial metabolites and colorectal cancer. [(accessed on 18 August 2020)];Nat. Rev. Microbiol. 2014 12:661–672. doi: 10.1113/expphysiol.2006.033837. Available online: www.nature.com/reviews/micro. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
