

Comparative Molecular Dynamics Investigation of the Electromotile Hearing Protein Prestin
- PMID: 34361083
- PMCID: PMC8347359
- DOI: 10.3390/ijms22158318
Comparative Molecular Dynamics Investigation of the Electromotile Hearing Protein Prestin
Abstract
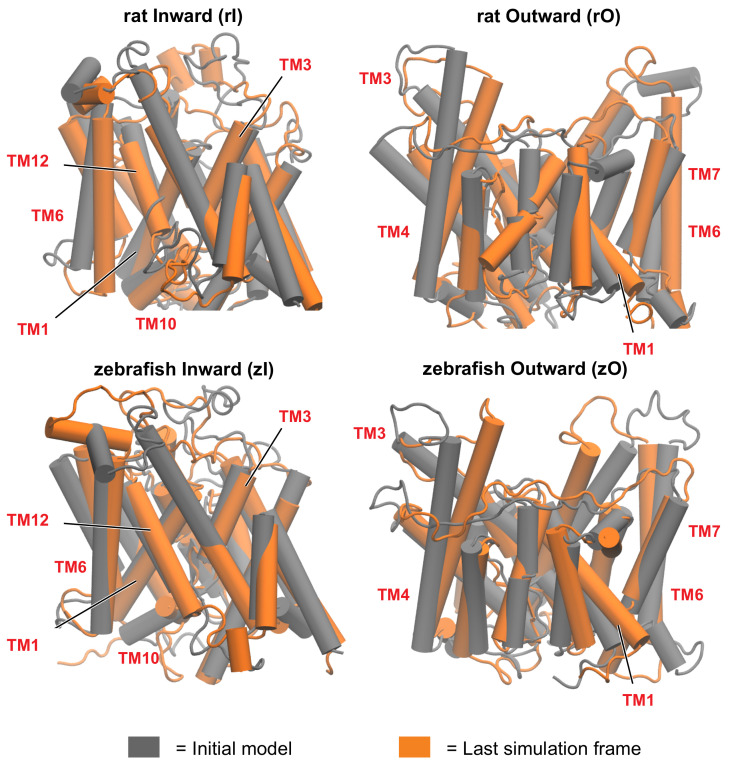
The mammalian protein prestin is expressed in the lateral membrane wall of the cochlear hair outer cells and is responsible for the electromotile response of the basolateral membrane, following hyperpolarisation or depolarisation of the cells. Its impairment marks the onset of severe diseases, like non-syndromic deafness. Several studies have pointed out possible key roles of residues located in the Transmembrane Domain (TMD) that differentiate mammalian prestins as incomplete transporters from the other proteins belonging to the same solute-carrier (SLC) superfamily, which are classified as complete transporters. Here, we exploit the homology of a prototypical incomplete transporter (rat prestin, rPres) and a complete transporter (zebrafish prestin, zPres) with target structures in the outward open and inward open conformations. The resulting models are then embedded in a model membrane and investigated via a rigorous molecular dynamics simulation protocol. The resulting trajectories are analyzed to obtain quantitative descriptors of the equilibration phase and to assess a structural comparison between proteins in different states, and between different proteins in the same state. Our study clearly identifies a network of key residues at the interface between the gate and the core domains of prestin that might be responsible for the conformational change observed in complete transporters and hindered in incomplete transporters. In addition, we study the pathway of Cl- ions in the presence of an applied electric field towards their putative binding site in the gate domain. Based on our simulations, we propose a tilt and shift mechanism of the helices surrounding the ion binding cavity as the working principle of the reported conformational changes in complete transporters.
Keywords: Non Linear Capacitance (NLC); SLC transporters; molecular dynamics simulations; prestin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures









References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
