

Mycobacterium tuberculosis VapC4 toxin engages small ORFs to initiate an integrated oxidative and copper stress response
- PMID: 34362841
- PMCID: PMC8364209
- DOI: 10.1073/pnas.2022136118
Mycobacterium tuberculosis VapC4 toxin engages small ORFs to initiate an integrated oxidative and copper stress response
Abstract
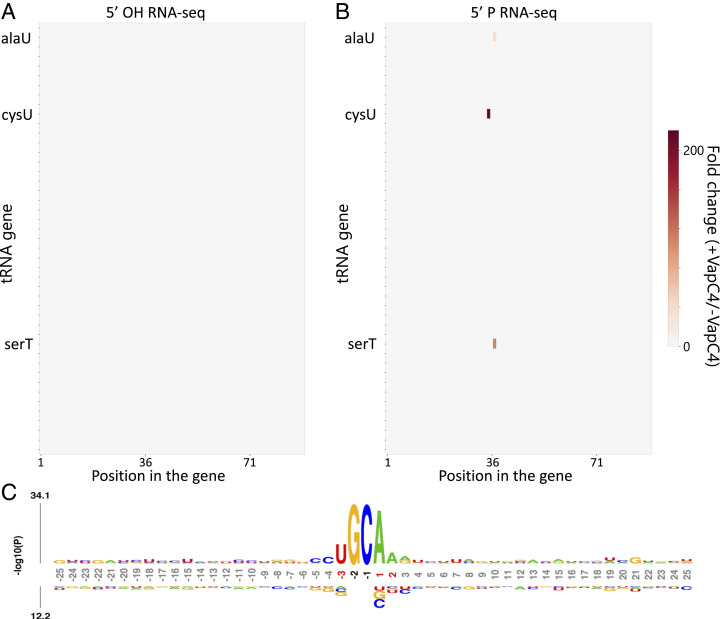
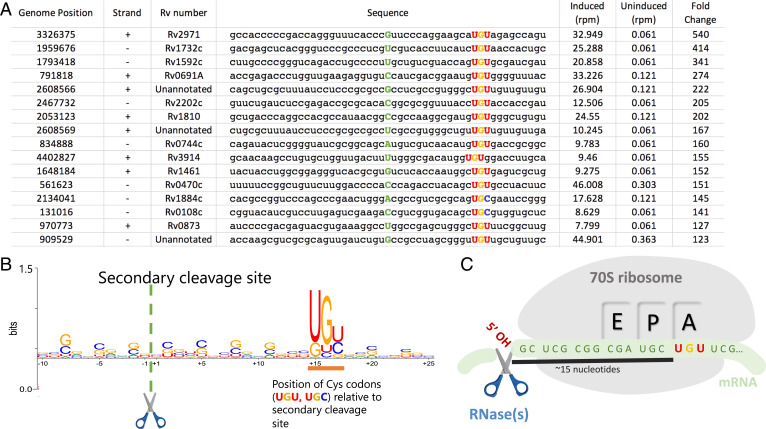
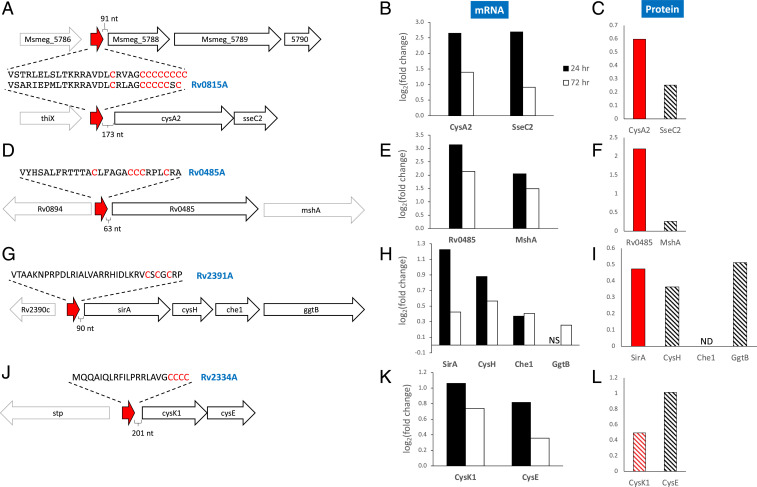
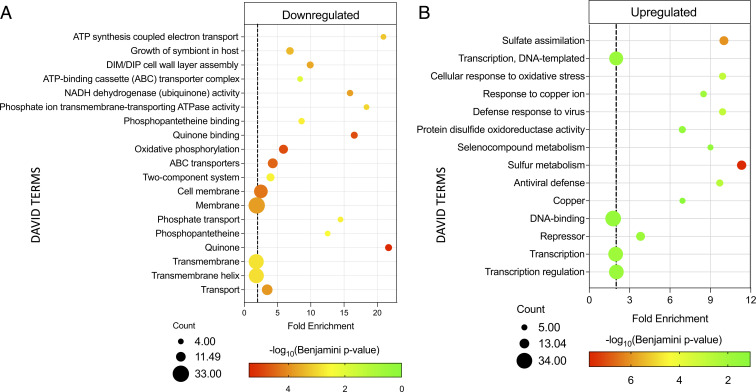
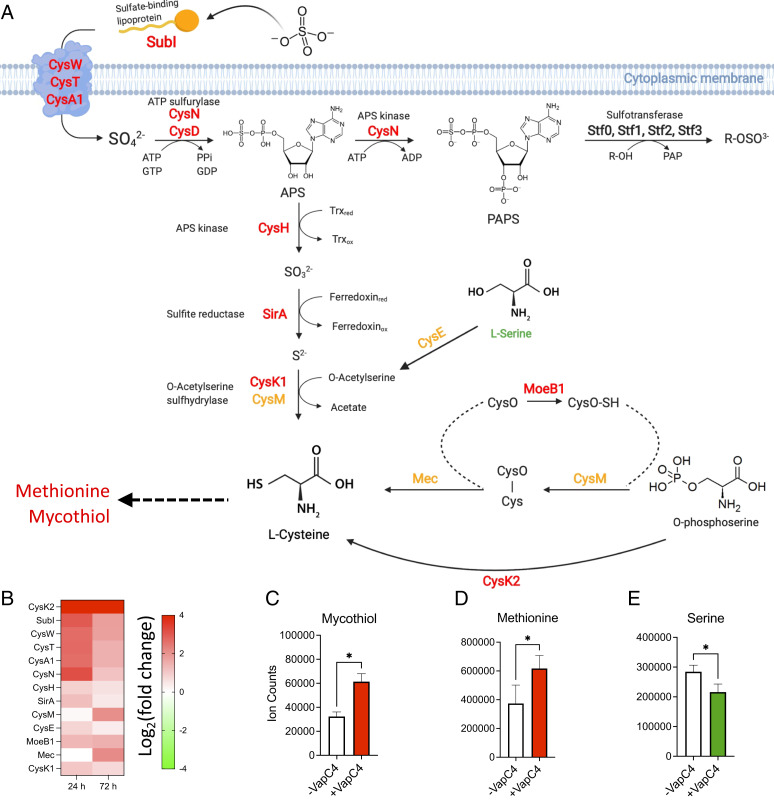
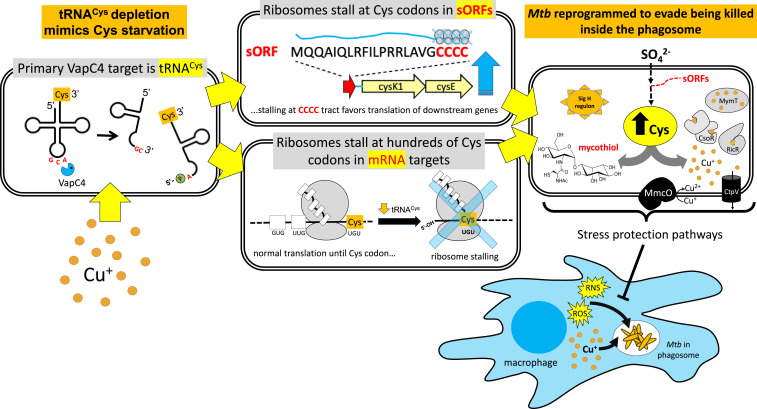
The Mycobacterium tuberculosis (Mtb) VapBC4 toxin-antitoxin system is essential for the establishment of Mtb infection. Using a multitier, systems-level approach, we uncovered the sequential molecular events triggered by the VapC4 toxin that activate a circumscribed set of critical stress survival pathways which undoubtedly underlie Mtb virulence. VapC4 exclusively inactivated the sole transfer RNACys (tRNACys) through cleavage at a single site within the anticodon sequence. Depletion of the pool of tRNACys led to ribosome stalling at Cys codons within actively translating messenger RNAs. Genome mapping of these Cys-stalled ribosomes unexpectedly uncovered several unannotated Cys-containing open reading frames (ORFs). Four of these are small ORFs (sORFs) encoding Cys-rich proteins of fewer than 50 amino acids that function as Cys-responsive attenuators that engage ribosome stalling at tracts of Cys codons to control translation of downstream genes. Thus, VapC4 mimics a state of Cys starvation, which then activates Cys attenuation at sORFs to globally redirect metabolism toward the synthesis of free Cys. The resulting newly enriched pool of Cys feeds into the synthesis of mycothiol, the glutathione counterpart in this pathogen that is responsible for maintaining cellular redox homeostasis during oxidative stress, as well as into a circumscribed subset of cellular pathways that enable cells to defend against oxidative and copper stresses characteristically endured by Mtb within macrophages. Our ability to pinpoint activation or down-regulation of pathways that collectively align with Mtb virulence-associated stress responses and the nonreplicating persistent state brings to light a direct and vital role for the VapC4 toxin in mediating these critical pathways.
Keywords: RNA-seq; mass spectrometry; mycothiol; protein translation; sulfur assimilation.
Conflict of interest statement
The authors declare no competing interest.
Figures



References
-
- Harms A., Brodersen D. E., Mitarai N., Gerdes K., Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 70, 768–784. (2018). - PubMed
-
- Arcus V. L., McKenzie J. L., Robson J., Cook G. M., The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng. Des. Sel. 24, 33–40 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
