

TRIM31 facilitates K27-linked polyubiquitination of SYK to regulate antifungal immunity
- PMID: 34362877
- PMCID: PMC8342987
- DOI: 10.1038/s41392-021-00711-3
TRIM31 facilitates K27-linked polyubiquitination of SYK to regulate antifungal immunity
Abstract
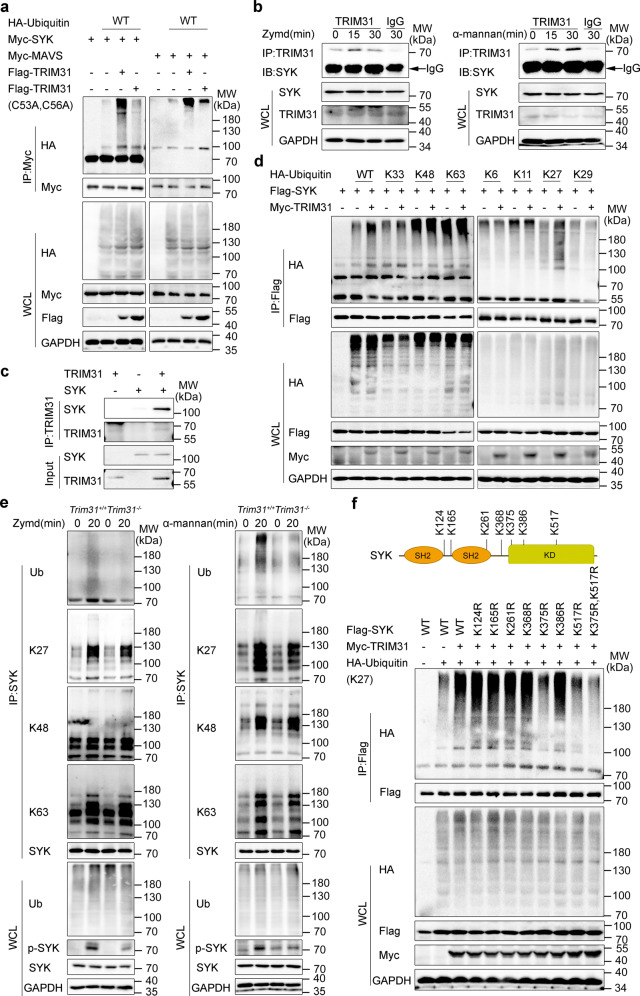
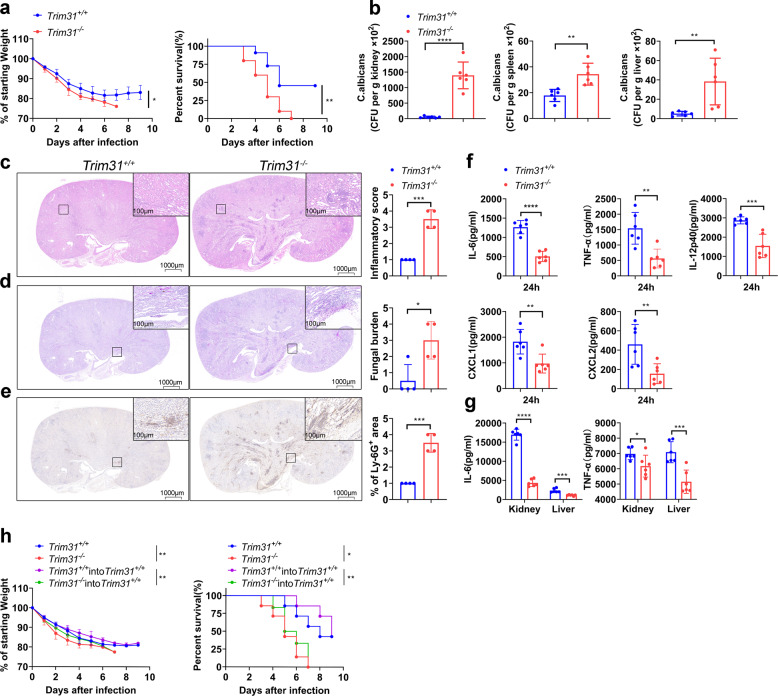
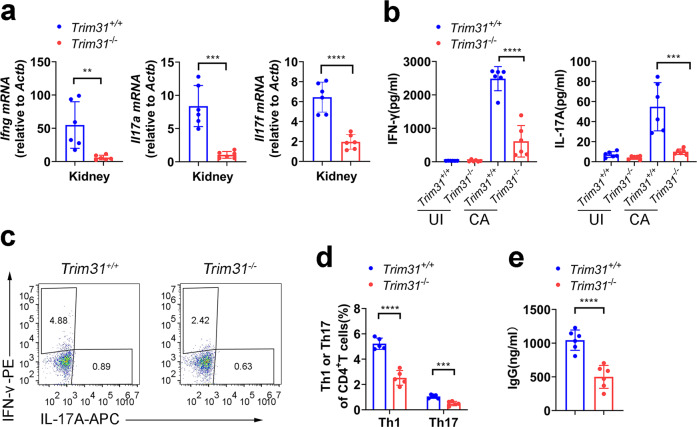
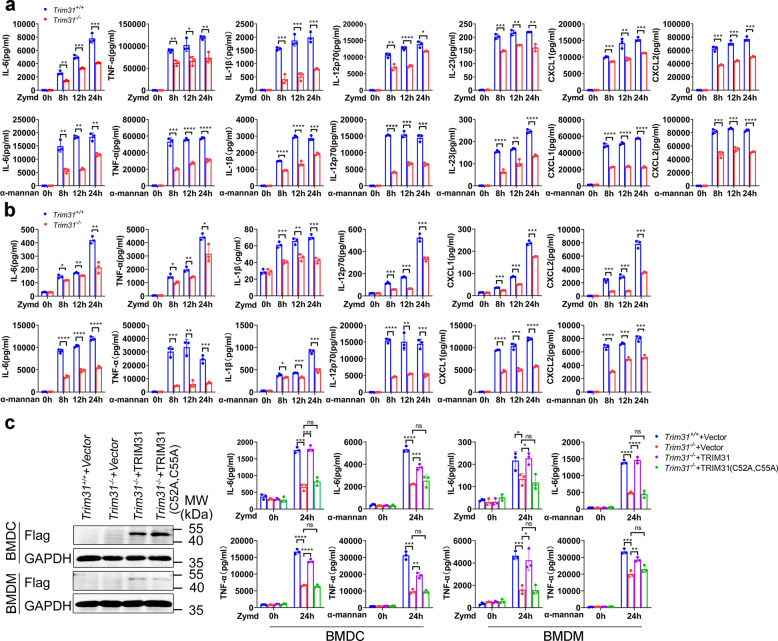
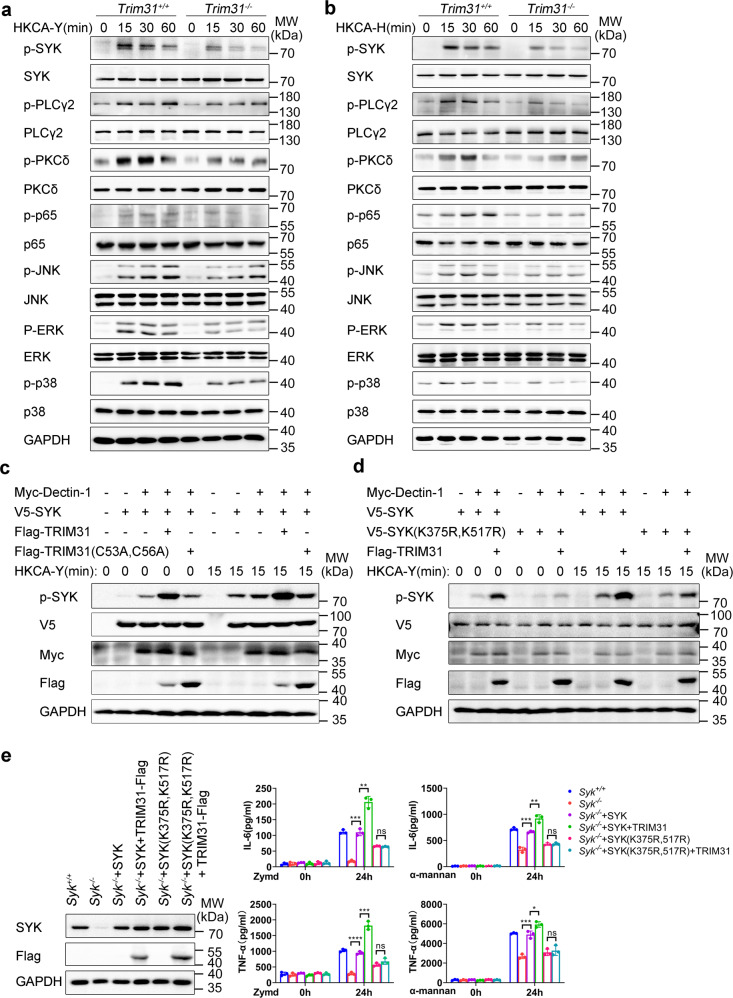
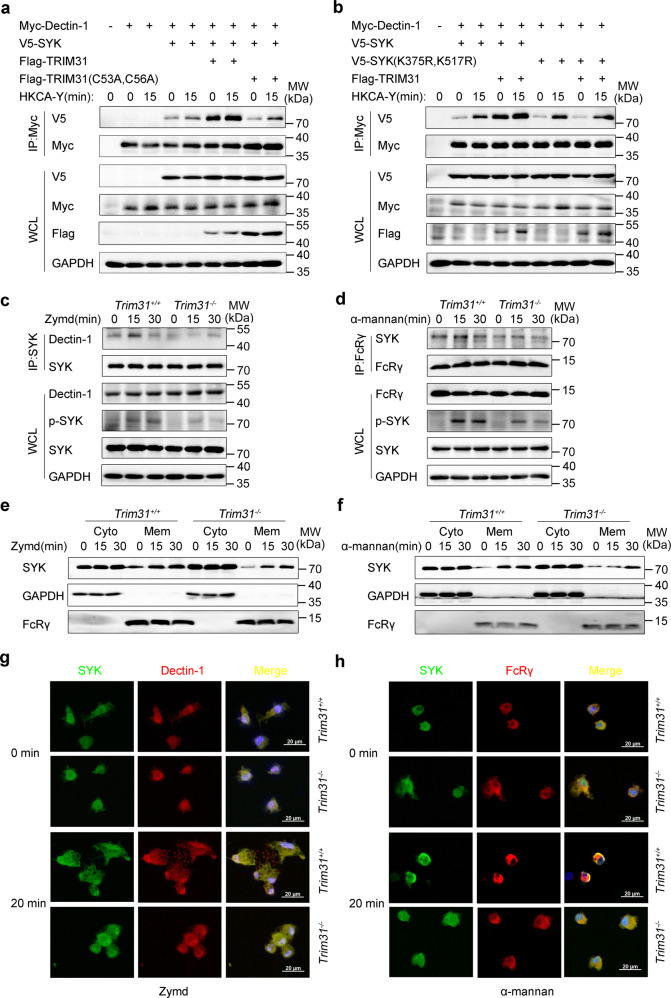
Spleen tyrosine kinase (SYK) is a non-receptor tyrosine kinase, which plays an essential role in both innate and adaptive immunity. However, the key molecular mechanisms that regulate SYK activity are poorly understood. Here we identified the E3 ligase TRIM31 as a crucial regulator of SYK activation. We found that TRIM31 interacted with SYK and catalyzed K27-linked polyubiquitination at Lys375 and Lys517 of SYK. This K27-linked polyubiquitination of SYK promoted its plasma membrane translocation and binding with the C-type lectin receptors (CLRs), and also prevented the interaction with the phosphatase SHP-1. Therefore, deficiency of Trim31 in bone marrow-derived dendritic cells (BMDCs) and macrophages (BMDMs) dampened SYK-mediated signaling and inhibited the secretion of proinflammatory cytokines and chemokines against the fungal pathogen Candida albicans infection. Trim31-/- mice were also more sensitive to C. albicans systemic infection than Trim31+/+ mice and exhibited reduced Th1 and Th17 responses. Overall, our study uncovered the pivotal role of TRIM31-mediated K27-linked polyubiquitination on SYK activation and highlighted the significance of TRIM31 in anti-C. albicans immunity.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
