

Post-translational modification of RAS proteins
- PMID: 34365229
- PMCID: PMC8649064
- DOI: 10.1016/j.sbi.2021.06.015
Post-translational modification of RAS proteins
Abstract
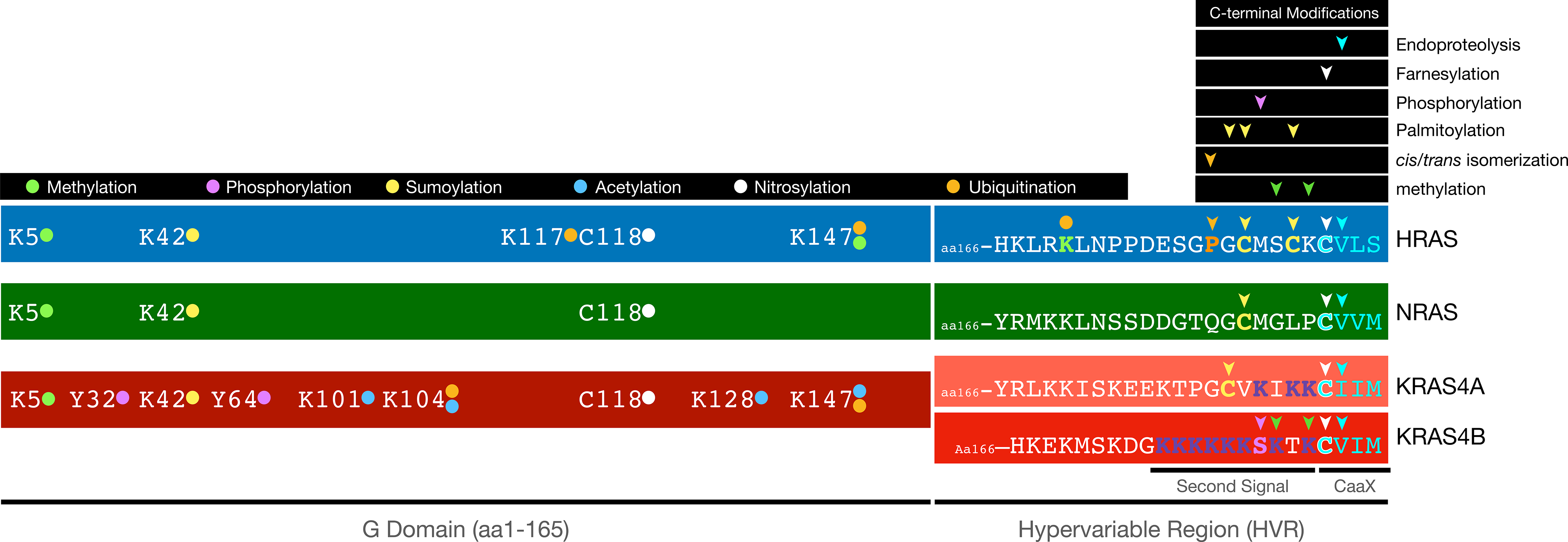
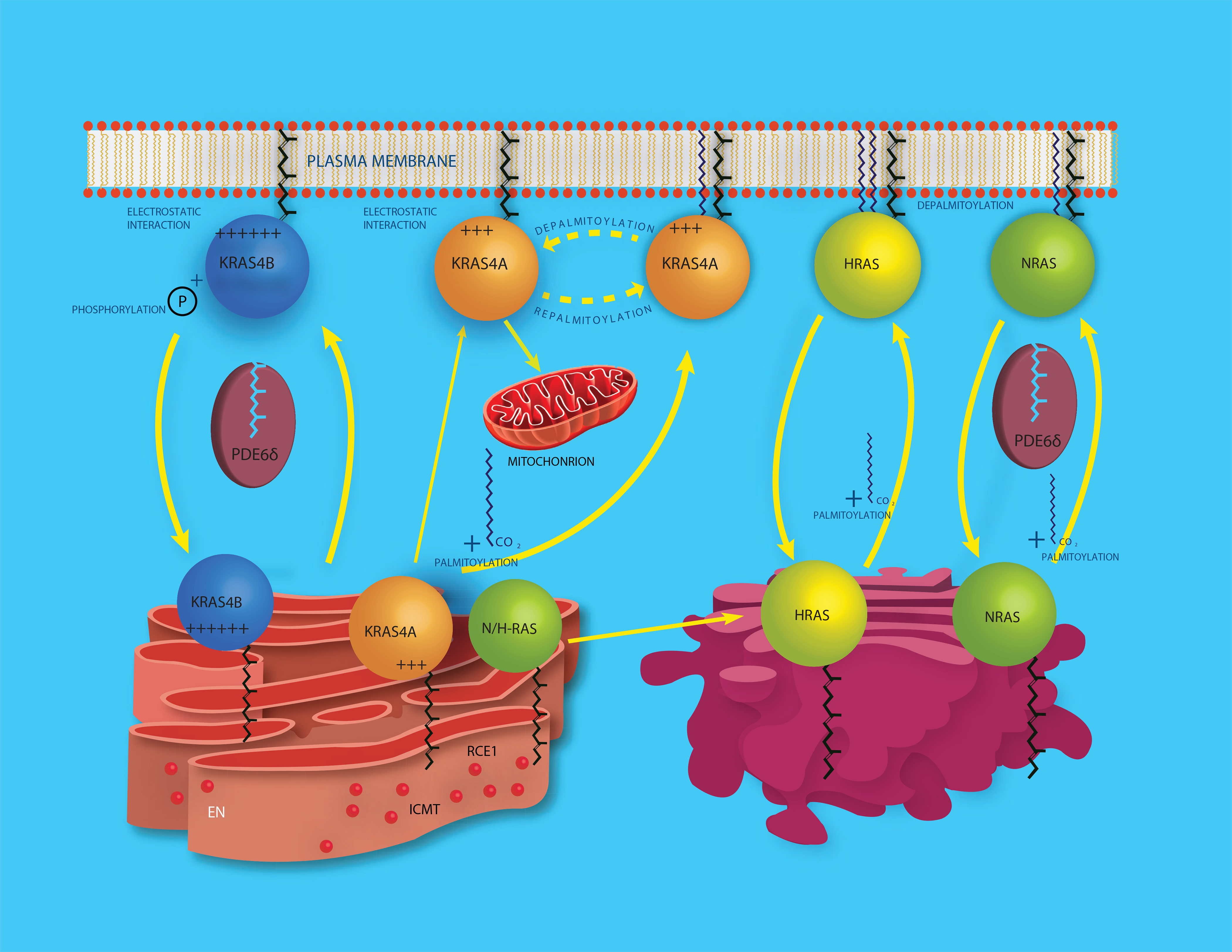
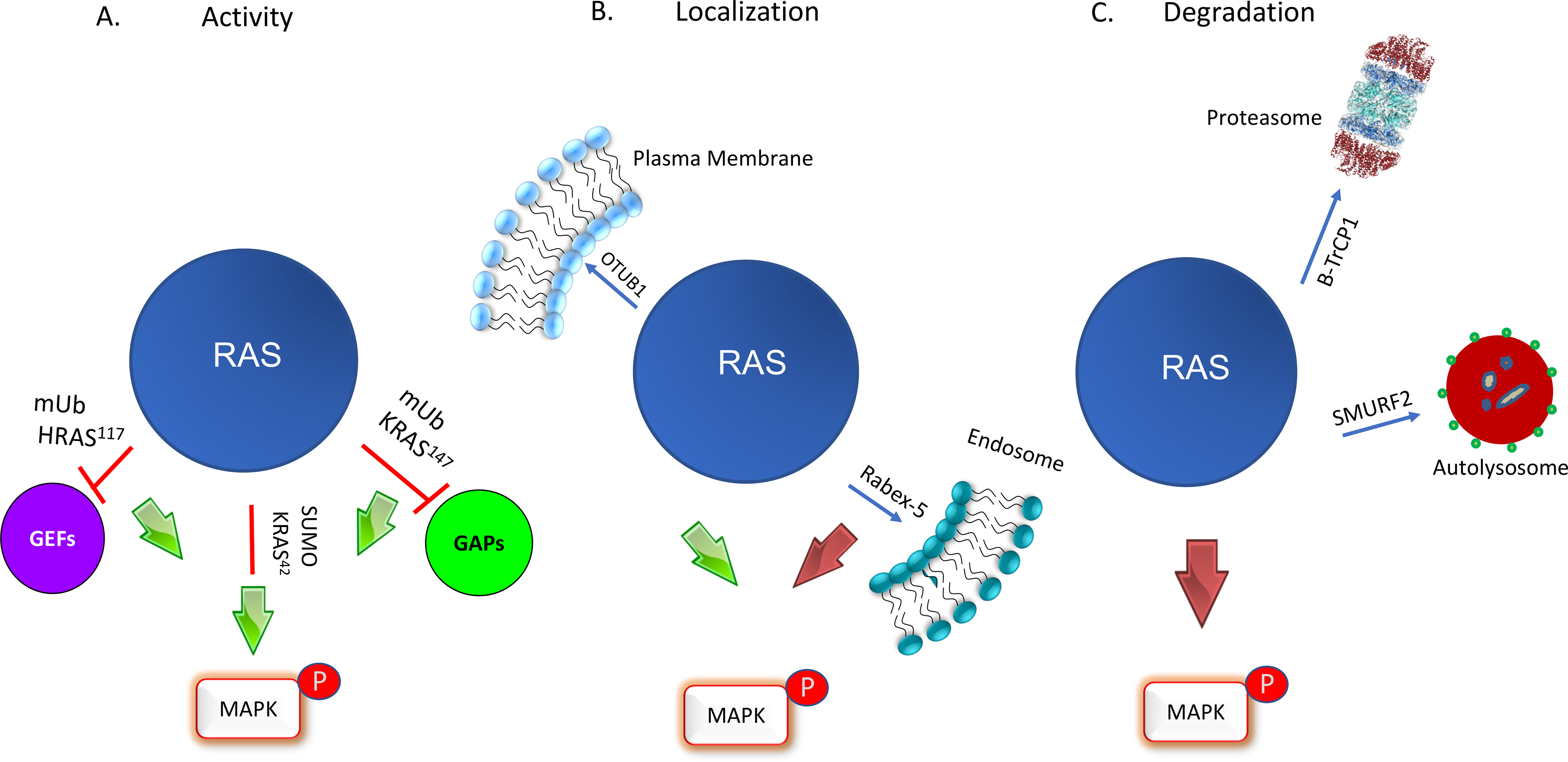
Mutations of RAS genes drive cancer more frequently than any other oncogene. RAS proteins integrate signals from a wide array of receptors and initiate downstream signaling through pathways that control cellular growth. RAS proteins are fundamentally binary molecular switches in which the off/on state is determined by the binding of GDP or GTP, respectively. As such, the intrinsic and regulated nucleotide-binding and hydrolytic properties of the RAS GTPase were historically believed to account for the entirety of the regulation of RAS signaling. However, it is increasingly clear that RAS proteins are also regulated by a vast array of post-translational modifications (PTMs). The current challenge is to understand what are the functional consequences of these modifications and which are physiologically relevant. Because PTMs are catalyzed by enzymes that may offer targets for drug discovery, the study of RAS PTMs has been a high priority for RAS biologists.
Keywords: Cancer; GTPase; Post-translational modification; RAS; Signaling.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Figures
References
-
- Wright LP, Philips MR: Thematic review series: lipid posttranslational modifications. CAAX modification and membrane targeting of Ras. J Lipid Res 2006, 47:883–891. - PubMed
-
- Buhaescu I, Izzedine H: Mevalonate pathway: a review of clinical and therapeutical implications. Clin Biochem 2007, 40:575–584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
