

A cascade of transcriptional repression determines sexual commitment and development in Plasmodium falciparum
- PMID: 34365503
- PMCID: PMC8450074
- DOI: 10.1093/nar/gkab683
A cascade of transcriptional repression determines sexual commitment and development in Plasmodium falciparum
Abstract
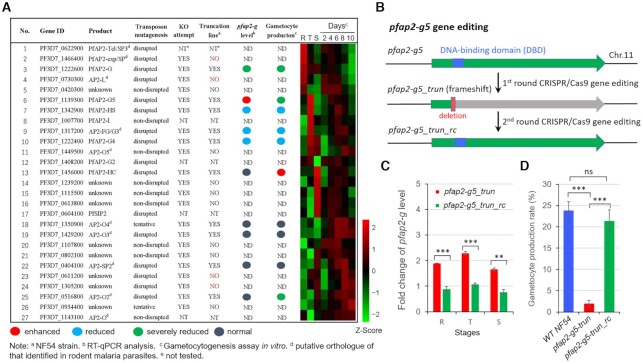
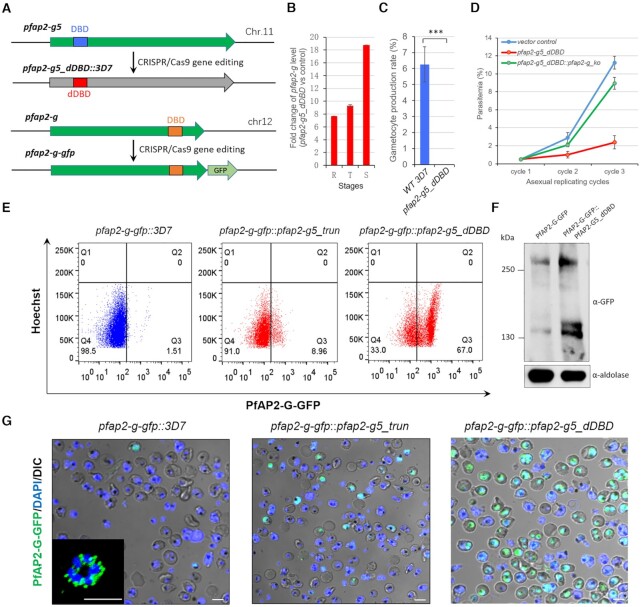
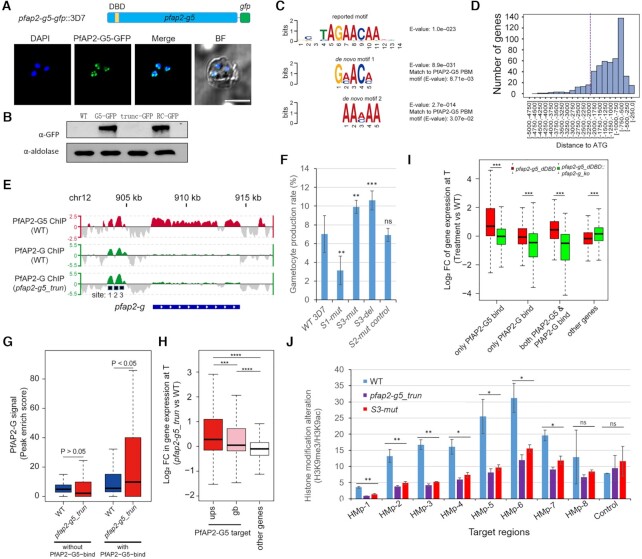
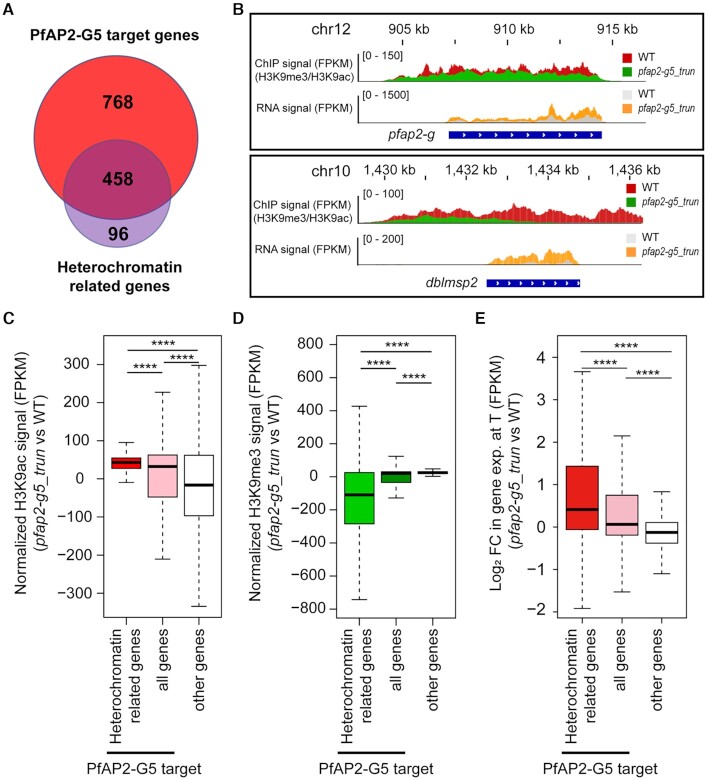
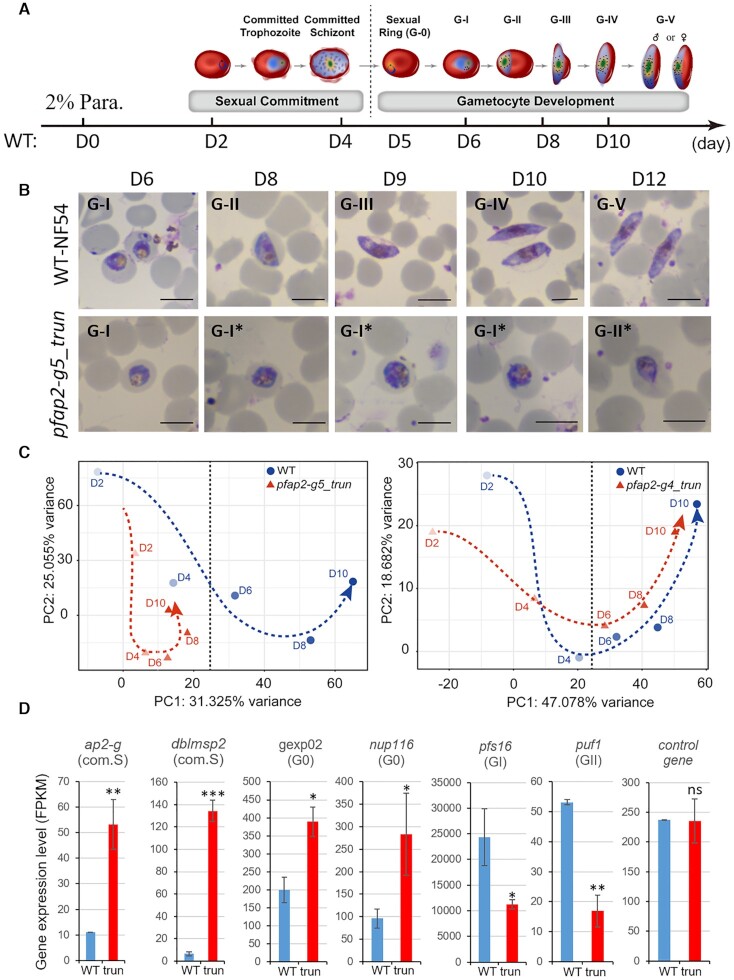
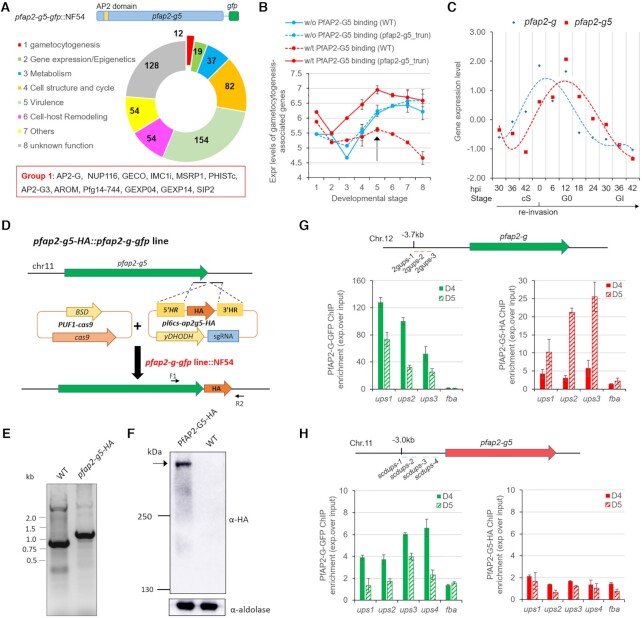
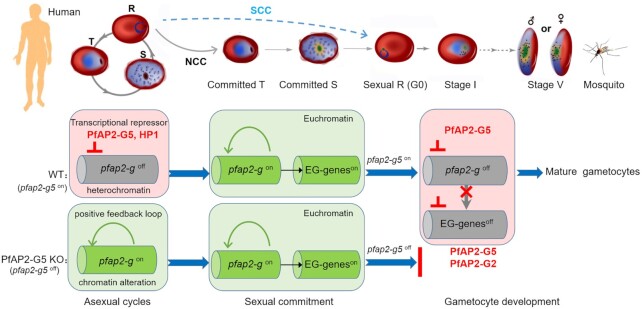
Gametocytogenesis, the process by which malaria parasites produce sexual forms that can infect mosquitoes, is essential for the transmission of malaria. A transcriptional switch of the pfap2-g gene triggers sexual commitment, but how the complex multi-step process is precisely programed remains largely unknown. Here, by systematic functional screening of a panel of ApiAP2 transcription factors, we identify six new ApiAP2 members associated with gametocytogenesis in Plasmodium falciparum. Among these, PfAP2-G5 (PF3D7_1139300) was found to be indispensable for gametocytogenesis. This factor suppresses the transcriptional activity of the pfap2-g gene via binding to both the upstream region and exonic gene body, the latter is linked to the maintenance of local heterochromatin structure, thereby preventing initiation of sexual commitment. Removal of this repressive effect through pfap2-g5 knockout disrupts the asexual replication cycle and promotes sexual commitment accompanied by upregulation of pfap2-g expression. However, the gametocytes produced fail to mature fully. Further analyses show that PfAP2-G5 is essential for gametocyte maturation, and causes the down-regulation of pfap2-g and a set of early gametocyte genes activated by PfAP2-G prior to gametocyte development. Collectively, our findings reveal a regulation cascade of gametocyte production in malaria parasites, and provide a new target for transmission blocking interventions.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Josling G.A., Llinás M.. Sexual development in Plasmodium parasites: knowing when it's time to commit. Nat. Rev. Microbiol. 2015; 13:573–587. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
