

Functional Relationship of Arabidopsis AOXs and PTOX Revealed via Transgenic Analysis
- PMID: 34367216
- PMCID: PMC8336870
- DOI: 10.3389/fpls.2021.692847
Functional Relationship of Arabidopsis AOXs and PTOX Revealed via Transgenic Analysis
Abstract
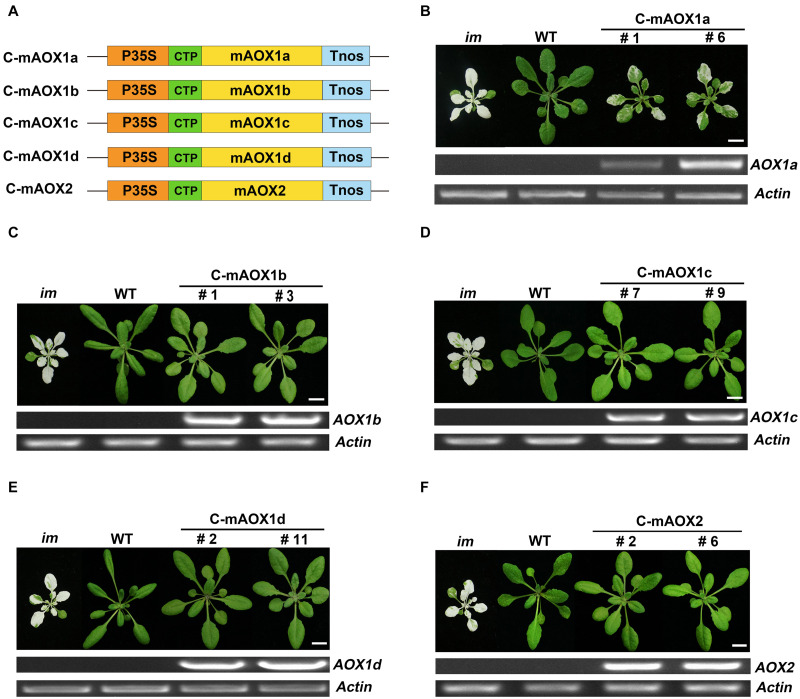
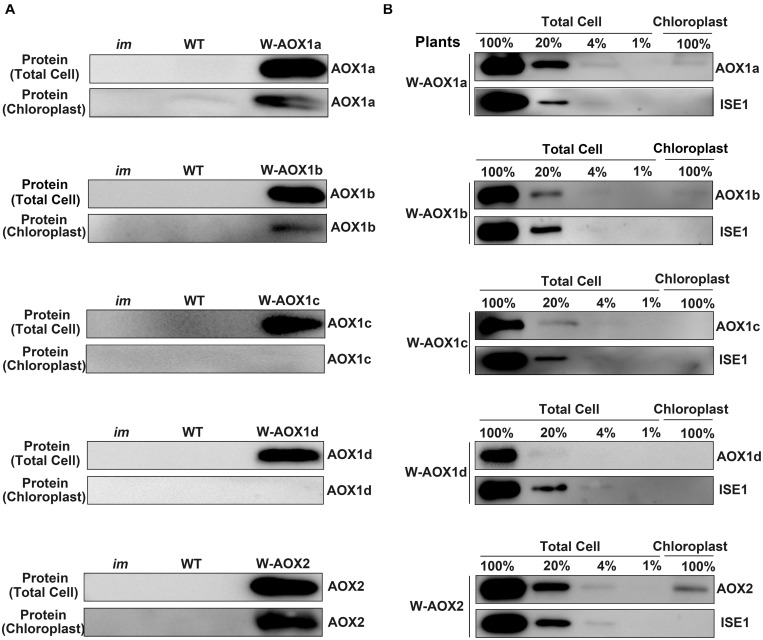
Alternative oxidase (AOX) and plastid terminal oxidase (PTOX) are terminal oxidases of electron transfer in mitochondria and chloroplasts, respectively. Here, taking advantage of the variegation phenotype of the Arabidopsis PTOX deficient mutant (im), we examined the functional relationship between PTOX and its five distantly related homologs (AOX1a, 1b, 1c, 1d, and AOX2). When engineered into chloroplasts, AOX1b, 1c, 1d, and AOX2 rescued the im defect, while AOX1a partially suppressed the mutant phenotype, indicating that AOXs could function as PQH2 oxidases. When the full length AOXs were overexpressed in im, only AOX1b and AOX2 rescued its variegation phenotype. In vivo fluorescence analysis of GFP-tagged AOXs and subcellular fractionation assays showed that AOX1b and AOX2 could partially enter chloroplasts while AOX1c and AOX1d were exclusively present in mitochondria. Surprisingly, the subcellular fractionation, but not the fluorescence analysis of GFP-tagged AOX1a, revealed that a small portion of AOX1a could sort into chloroplasts. We further fused and expressed the targeting peptides of AOXs with the mature form of PTOX in im individually; and found that targeting peptides of AOX1a, AOX1b, and AOX2, but not that of AOX1c or AOX1d, could direct PTOX into chloroplasts. It demonstrated that chloroplast-localized AOXs, but not mitochondria-localized AOXs, can functionally compensate for the PTOX deficiency in chloroplasts, providing a direct evidence for the functional relevance of AOX and PTOX, shedding light on the interaction between mitochondria and chloroplasts and the complex mechanisms of protein dual targeting in plant cells.
Keywords: alternative oxidase (AOX); chloroplasts; mitochondria; plastid terminal oxidase (PTOX); protein dual targeting; targeting peptide.
Copyright © 2021 Wang, Wang, Li, Song, Qin, Chang, Fu, Wang, Wang, Li, Hao, Xu and Fu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases