

Chromatin and transcriptomic profiling uncover dysregulation of the Tip60 HAT/HDAC2 epigenomic landscape in the neurodegenerative brain
- PMID: 34369292
- PMCID: PMC9336495
- DOI: 10.1080/15592294.2021.1959742
Chromatin and transcriptomic profiling uncover dysregulation of the Tip60 HAT/HDAC2 epigenomic landscape in the neurodegenerative brain
Abstract
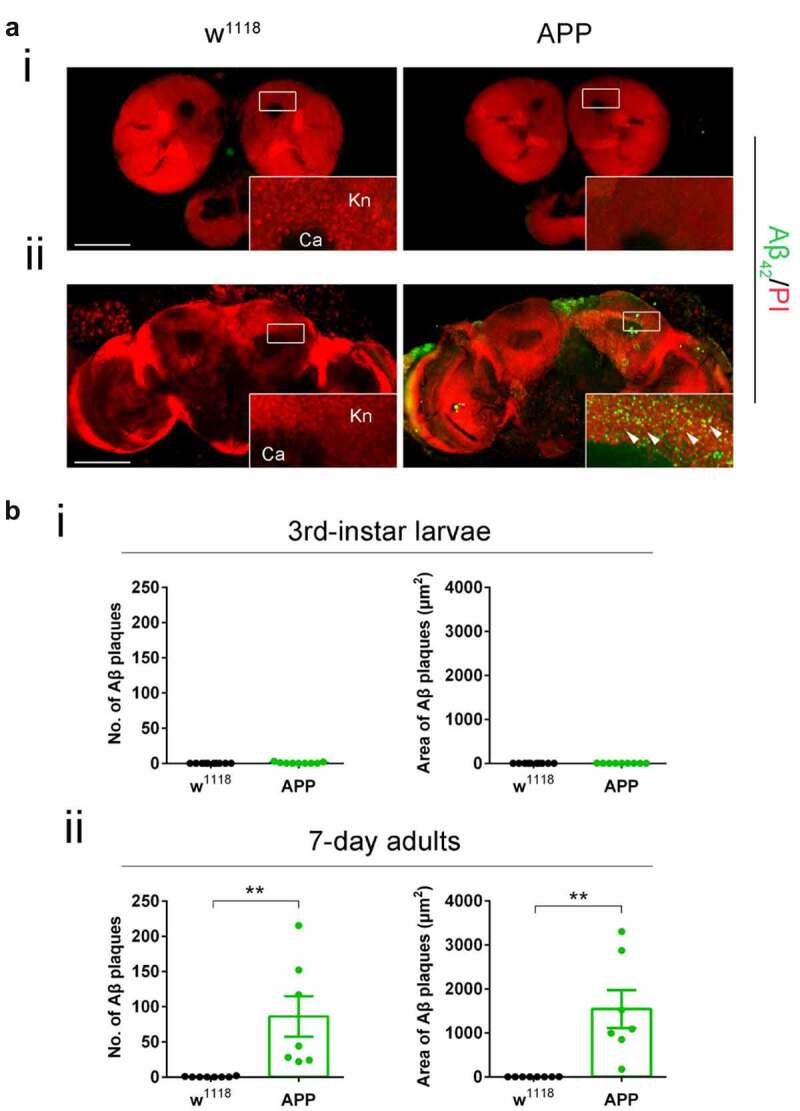
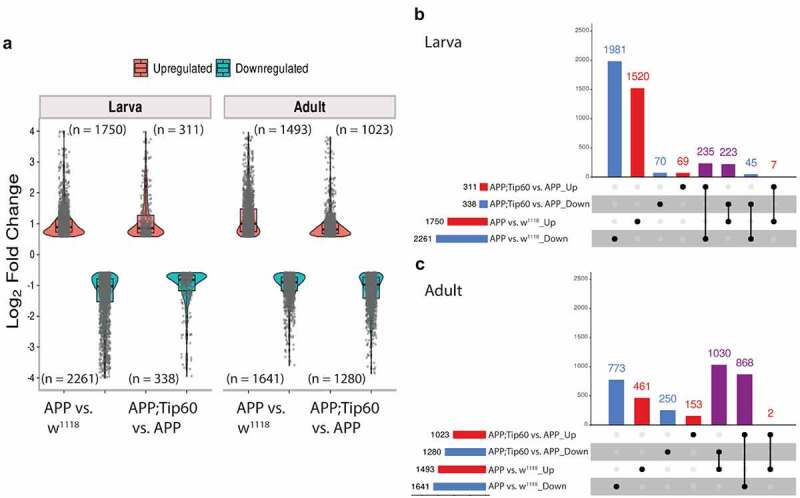
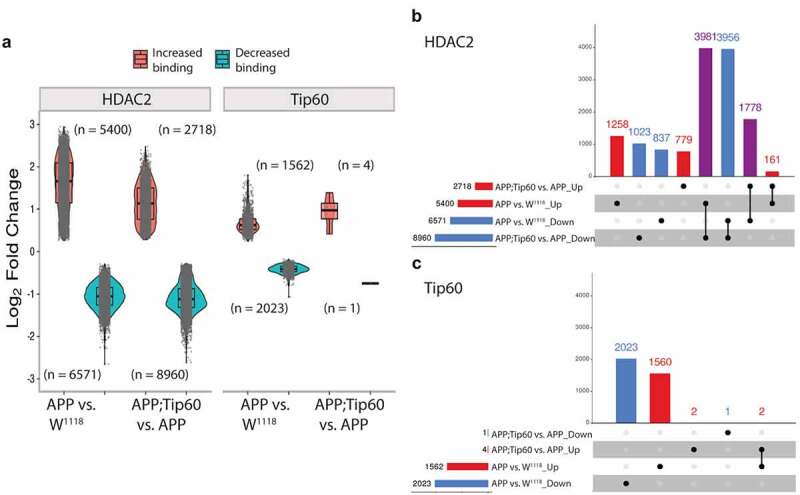
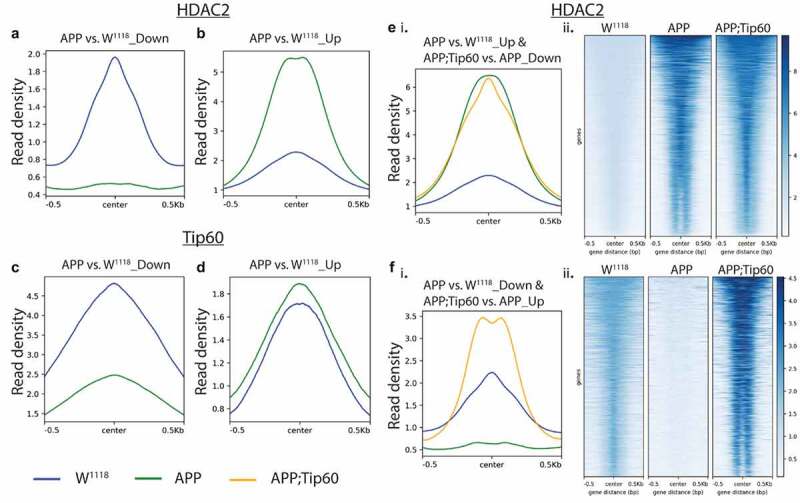
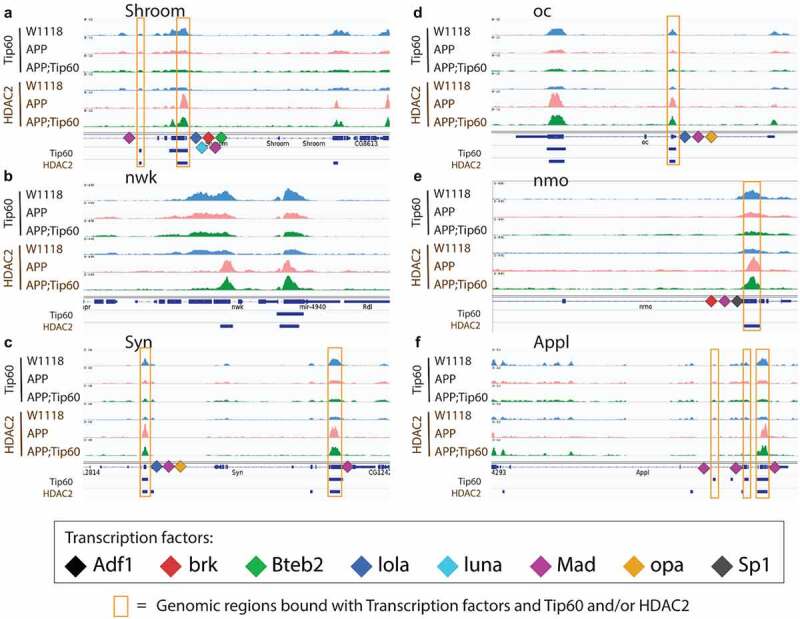
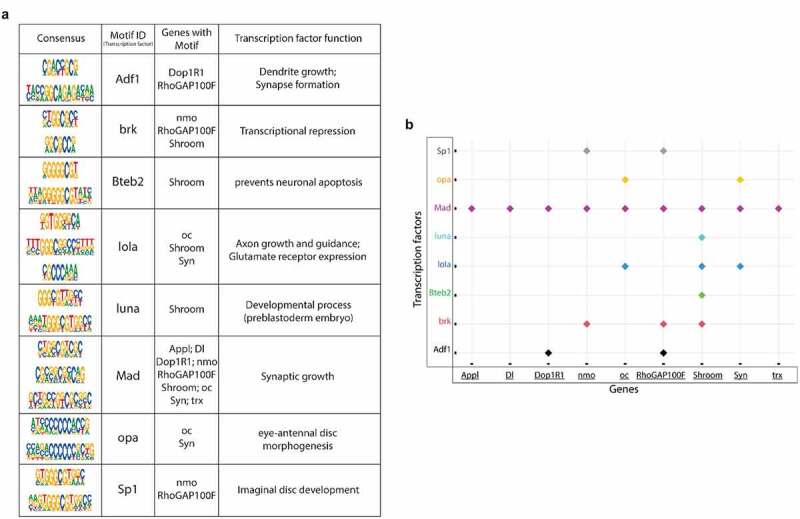
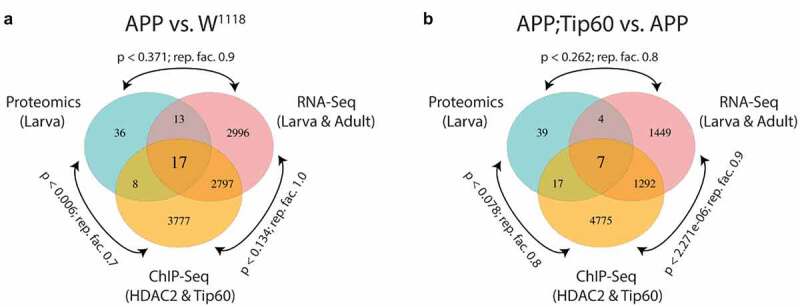
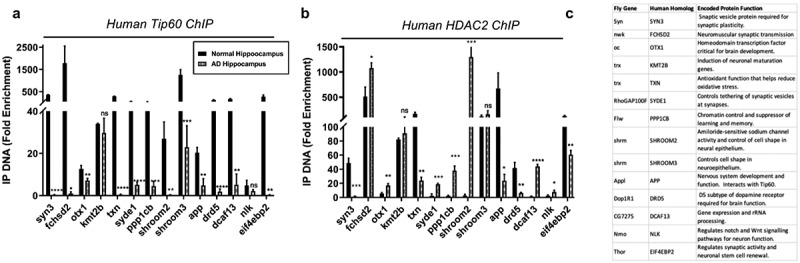
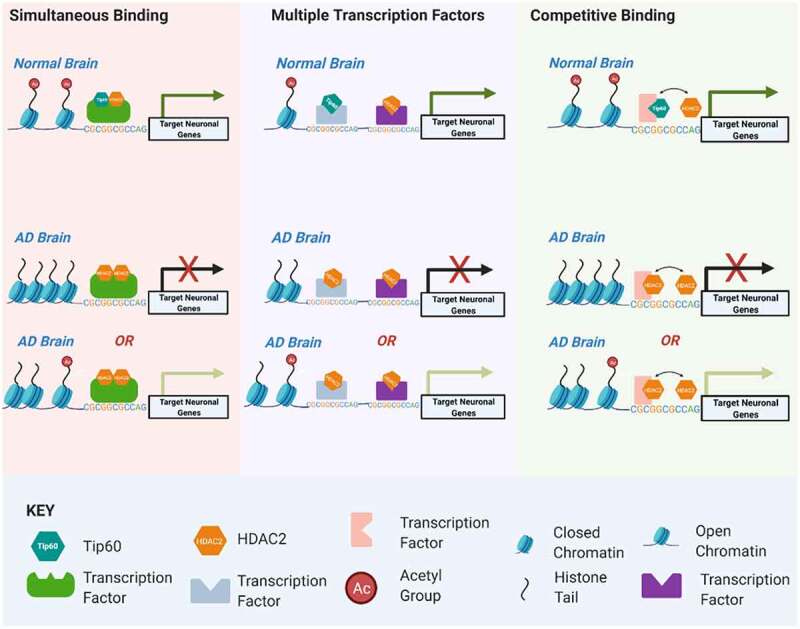
Disruption of histone acetylation-mediated gene control is a critical step in Alzheimer's Disease (AD), yet chromatin analysis of antagonistic histone acetyltransferases (HATs) and histone deacetylases (HDACs) causing these alterations remains uncharacterized. We report the first Tip60 HAT versus HDAC2 chromatin (ChIP-seq) and transcriptional (RNA-seq) profiling study in Drosophila melanogaster brains that model early human AD. We find Tip60 and HDAC2 predominantly recruited to identical neuronal genes. Moreover, AD brains exhibit robust genome-wide early alterations that include enhanced HDAC2 and reduced Tip60 binding and transcriptional dysregulation. Orthologous human genes to co-Tip60/HDAC2 D. melanogaster neural targets exhibit conserved disruption patterns in AD patient hippocampi. Notably, we discovered distinct transcription factor binding sites close or within Tip60/HDAC2 co-peaks in neuronal genes, implicating them in coenzyme recruitment. Increased Tip60 protects against transcriptional dysregulation and enhanced HDAC2 enrichment genome-wide. We advocate Tip60 HAT/HDAC2 mediated epigenetic neuronal gene disruption as a genome-wide initial causal event in AD.
Keywords: Alzheimer’s disease; Amyloid precursor protein; KAT5; histone acetylation; sequencing.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures

References
-
- Masters CL, Bateman R, Blennow K, et al. Alzheimer’s disease. Nat Rev Dis Primers. 2015;1(1):15056. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous