

Structural Characterization of the Reaction and Substrate Specificity Mechanisms of Pathogenic Fungal Acetyl-CoA Synthetases
- PMID: 34369755
- PMCID: PMC8383264
- DOI: 10.1021/acschembio.1c00484
Structural Characterization of the Reaction and Substrate Specificity Mechanisms of Pathogenic Fungal Acetyl-CoA Synthetases
Abstract
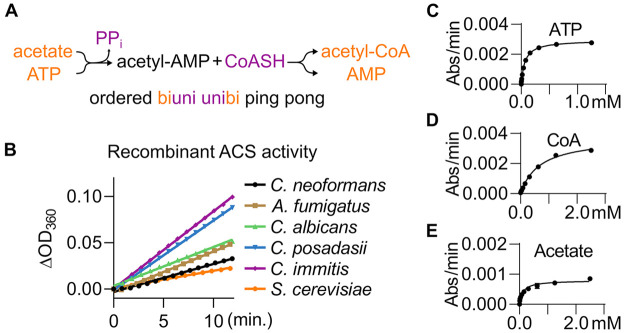
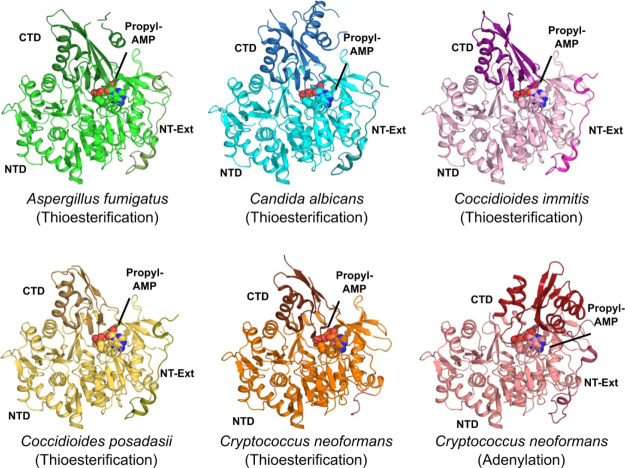
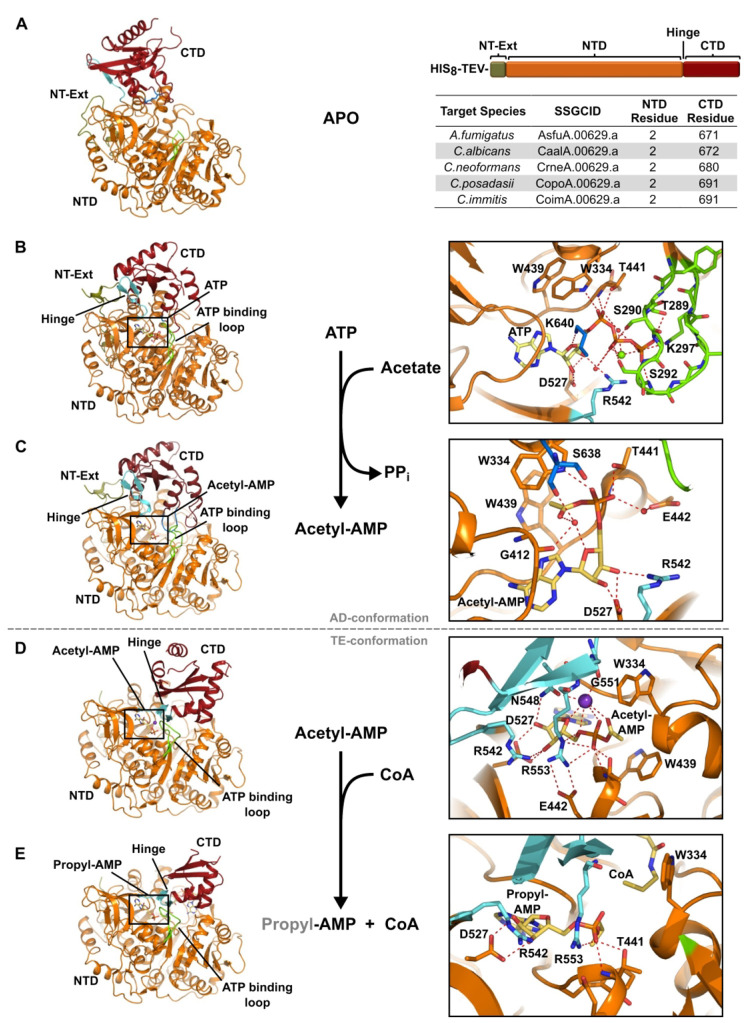
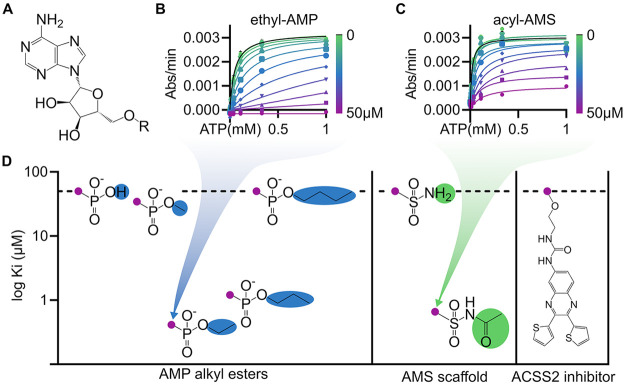
Acetyl CoA synthetases (ACSs) are
Conflict of interest statement
The authors declare no competing financial interest.
Figures
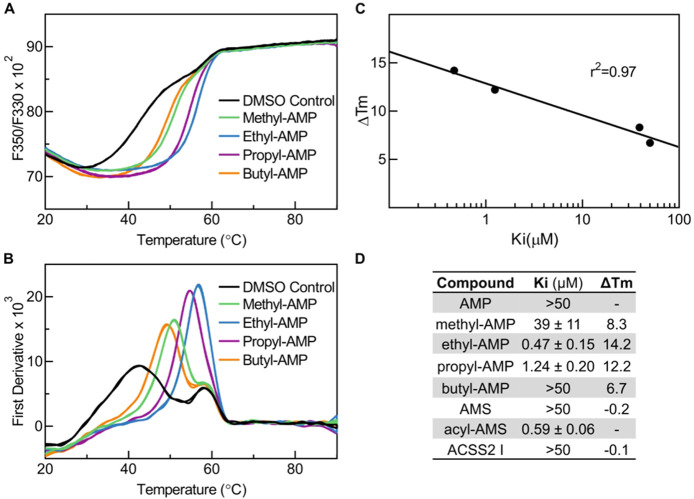
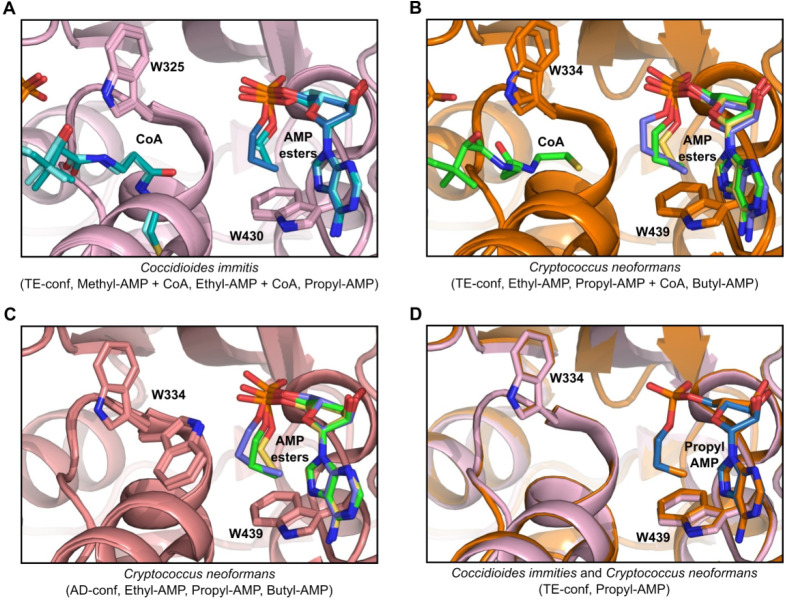
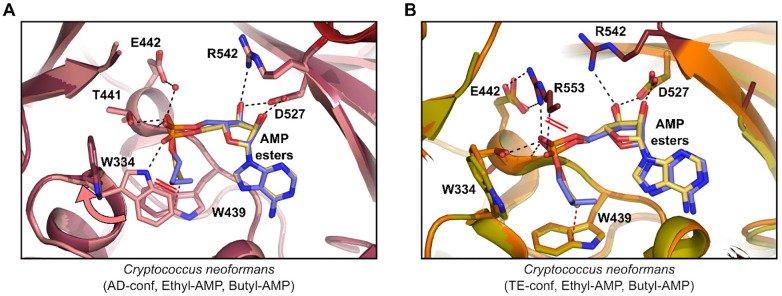
References
-
- Mashimo T.; Pichumani K.; Vemireddy V.; Hatanpaa K. J.; Singh D. K.; Sirasanagandla S.; Nannepaga S.; Piccirillo S. G.; Kovacs Z.; Foong C.; et al. (2014) Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell 159, 1603–1614. 10.1016/j.cell.2014.11.025. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
