

Ageing-associated effects of a long-term dietary modulation of four trace elements in mice
- PMID: 34371368
- PMCID: PMC8358688
- DOI: 10.1016/j.redox.2021.102083
Ageing-associated effects of a long-term dietary modulation of four trace elements in mice
Abstract
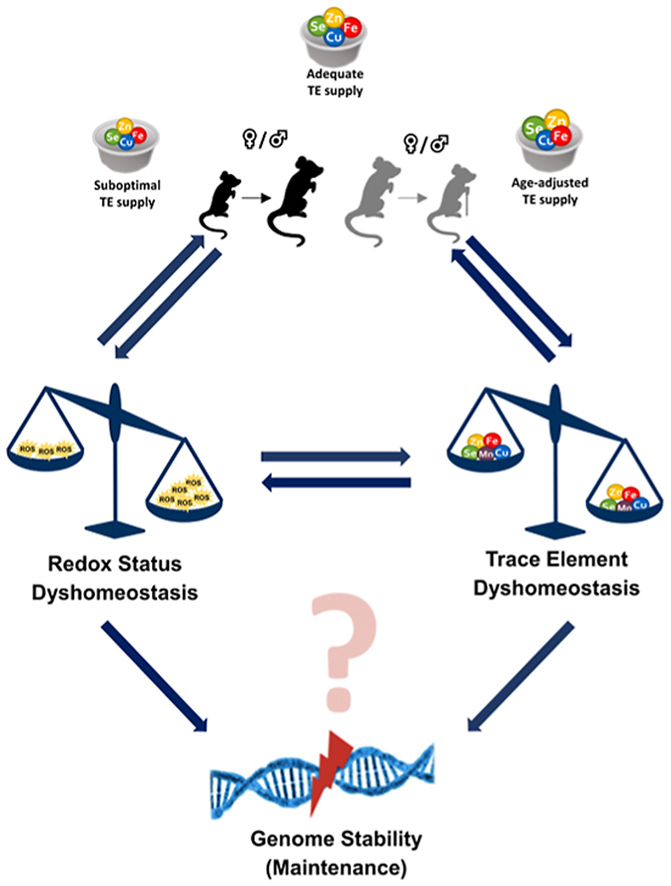
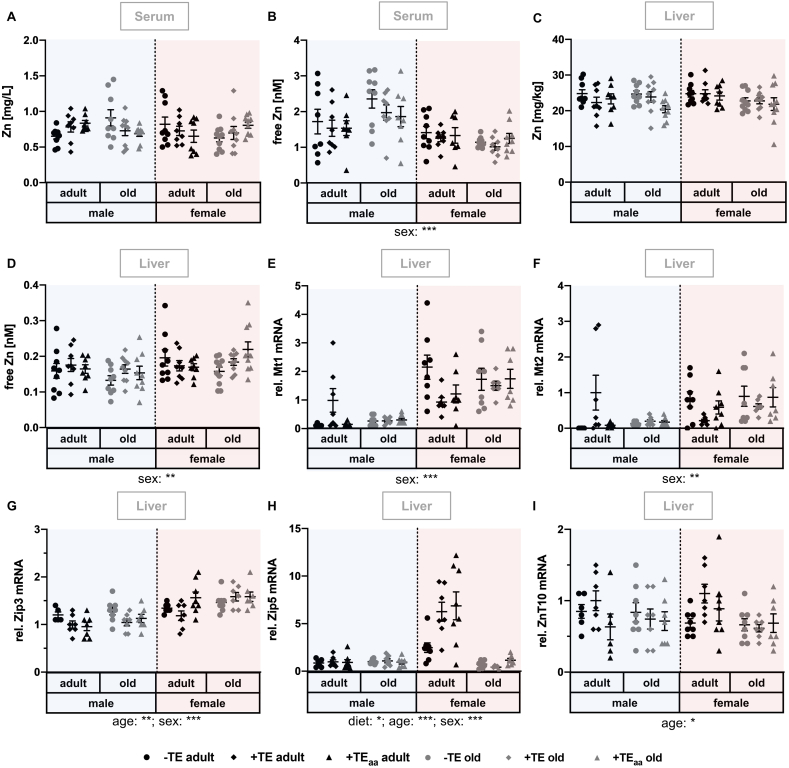
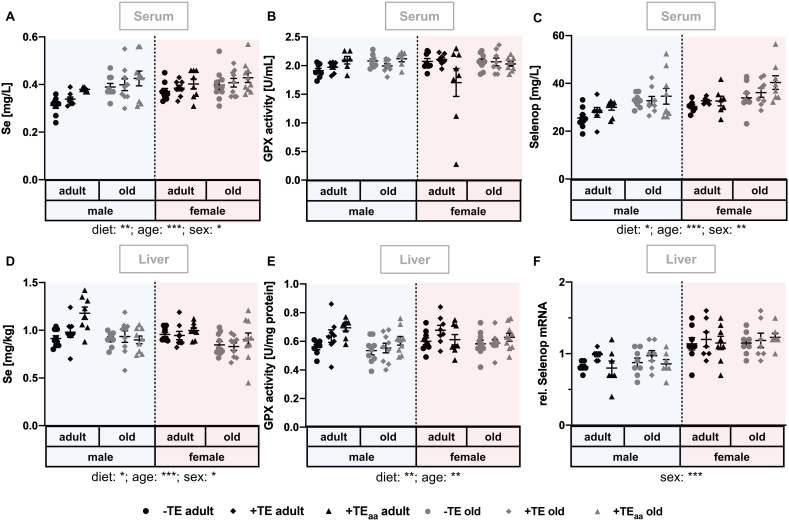
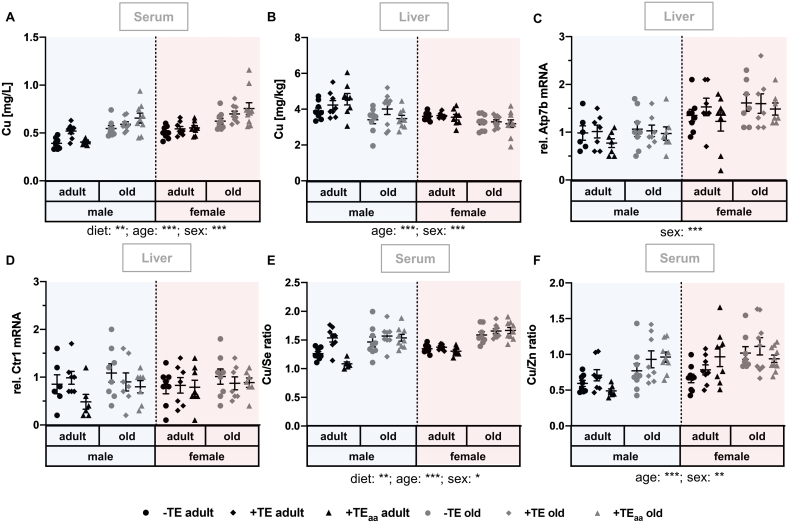
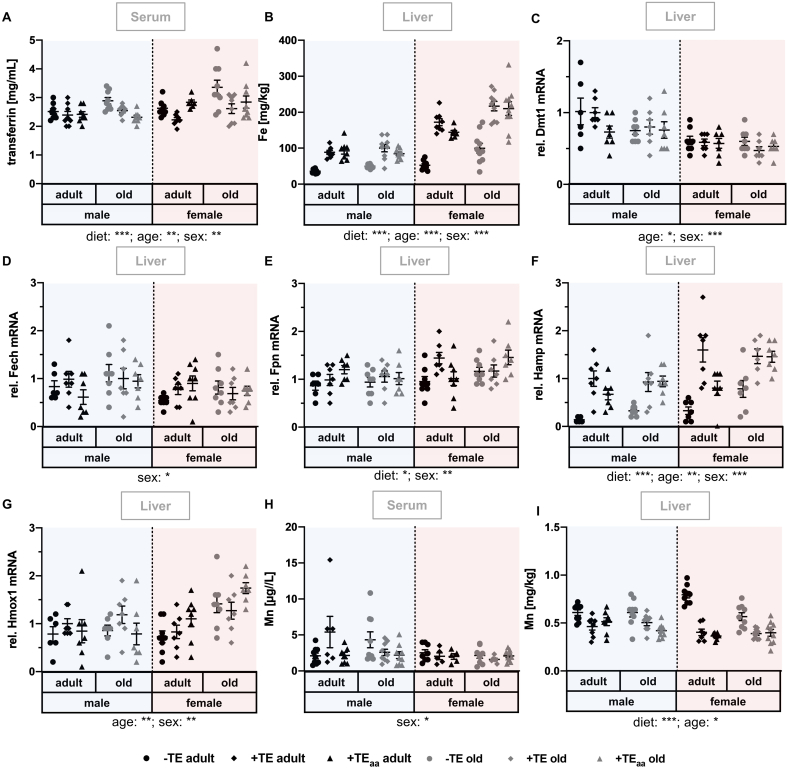
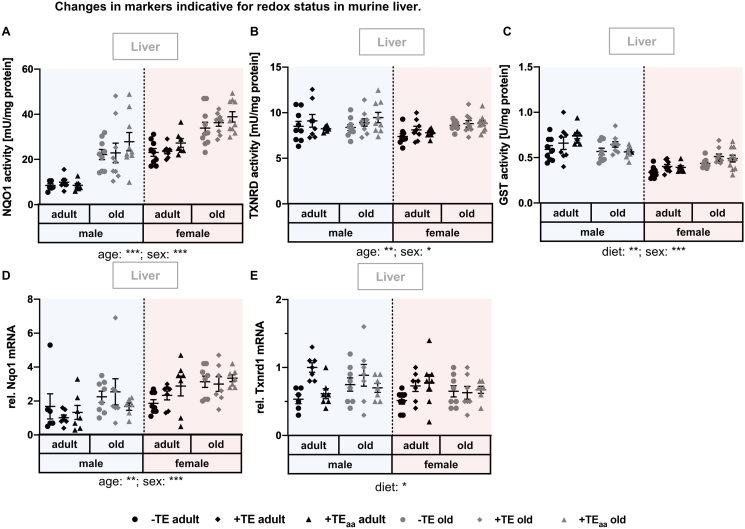
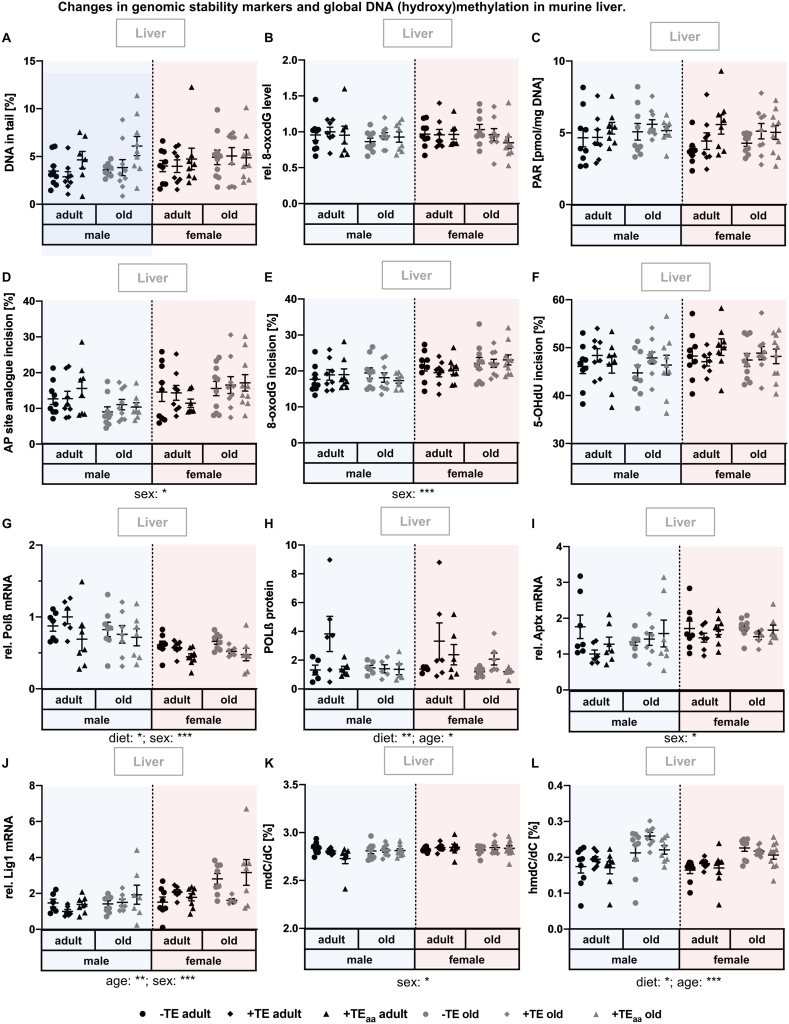
Trace elements (TEs) are essential for diverse processes maintaining body function and health status. The complex regulation of the TE homeostasis depends among others on age, sex, and nutritional status. If the TE homeostasis is disturbed, negative health consequences can result, e.g., caused by impaired redox homeostasis and genome stability maintenance. Based on age-related shifts in TEs which have been described in mice well-supplied with TEs, we aimed to understand effects of a long-term feeding with adequate or suboptimal amounts of four TEs in parallel. As an additional intervention, we studied mice which received an age-adapted diet with higher concentrations of selenium and zinc to counteract the age-related decline of both TEs. We conducted comprehensive analysis of diverse endpoints indicative for the TE and redox status, complemented by analysis of DNA (hydroxy)methylation and markers denoting genomic stability maintenance. TE concentrations showed age-specific alterations which were relatively stable and independent of their nutritional supply. In addition, hepatic DNA hydroxymethylation was significantly increased in the elderly mice and markers indicative for the redox status were modulated. The reduced nutritional supply with TEs inconsistently affected their status, with most severe effects regarding Fe deficiency. This may have contributed to the sex-specific differences observed in the alterations related to the redox status and DNA repair activity. Overall, our results highlight the complexity of factors impacting on the TE status and its physiological consequences. Alterations in TE supply, age, and sex proved to be important determinants that need to be taken into account when considering TE interventions for improving general health and supporting convalescence in the clinics.
Keywords: Ageing; Genome stability (maintenance); Redox status; Sex; Trace elements.
Copyright © 2021 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Bornhorst J., Kipp A.P., Haase H., Meyer S., Schwerdtle T. The crux of inept biomarkers for risks and benefits of trace elements. Trends Anal. Chem. 2018;104:183–190.
-
- WHO Trace elements in human nutrition. Report of a WHO expert committee. World Health Organ Tech Rep Ser. 1973;532:1–65. - PubMed
-
- Malbohan I.M., Fialova L. [Trace elements, human nutrition and health] Cas. Lek. Cesk. 1997;136(11):356–359. - PubMed
-
- Mertz W. The essential trace elements. Science. 1981;213(4514):1332–1338. - PubMed
-
- Livingstone C. Zinc: physiology, deficiency, and parenteral nutrition. Nutr. Clin. Pract. 2015;30(3):371–382. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
