

A genetically encoded tool for reconstituting synthetic modulatory neurotransmission and reconnect neural circuits in vivo
- PMID: 34373460
- PMCID: PMC8352926
- DOI: 10.1038/s41467-021-24690-9
A genetically encoded tool for reconstituting synthetic modulatory neurotransmission and reconnect neural circuits in vivo
Abstract
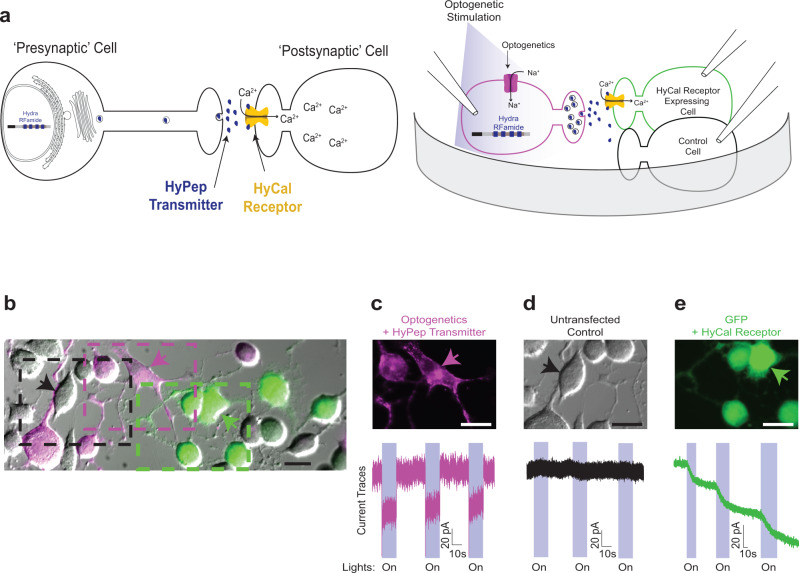
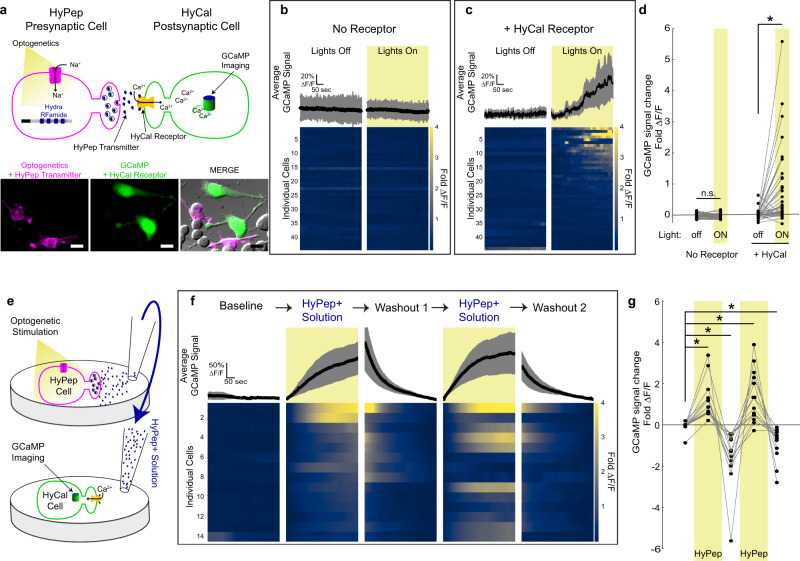
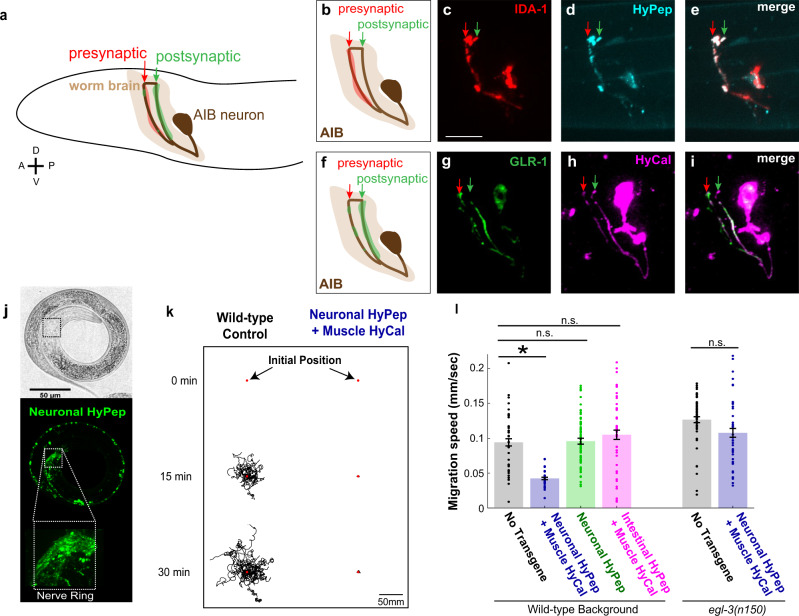
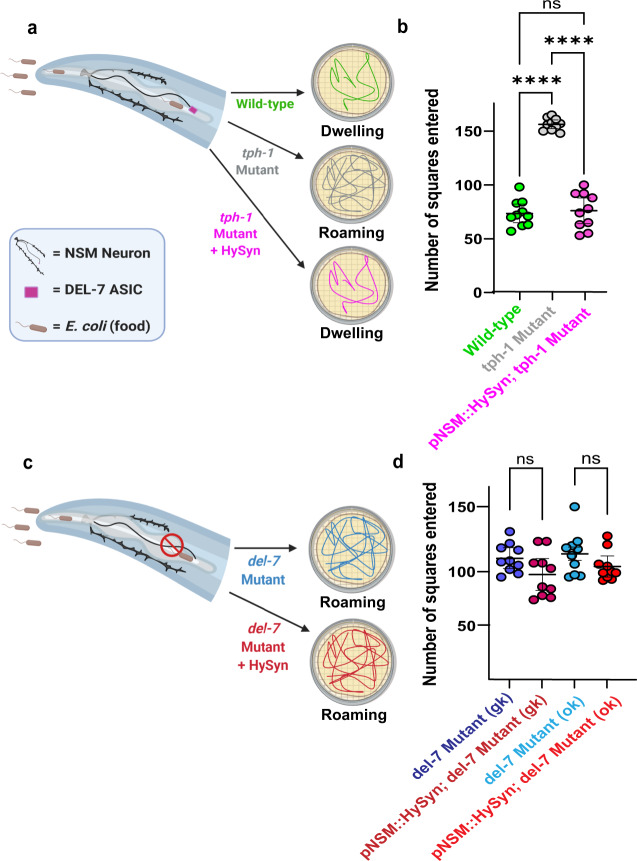
Chemogenetic and optogenetic tools have transformed the field of neuroscience by facilitating the examination and manipulation of existing circuits. Yet, the field lacks tools that enable rational rewiring of circuits via the creation or modification of synaptic relationships. Here we report the development of HySyn, a system designed to reconnect neural circuits in vivo by reconstituting synthetic modulatory neurotransmission. We demonstrate that genetically targeted expression of the two HySyn components, a Hydra-derived neuropeptide and its receptor, creates de novo neuromodulatory transmission in a mammalian neuronal tissue culture model and functionally rewires a behavioral circuit in vivo in the nematode Caenorhabditis elegans. HySyn can interface with existing optogenetic, chemogenetic and pharmacological approaches to functionally probe synaptic transmission, dissect neuropeptide signaling, or achieve targeted modulation of specific neural circuits and behaviors.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Illuminating neural circuits and behaviour in Caenorhabditis elegans with optogenetics.Philos Trans R Soc Lond B Biol Sci. 2015 Sep 19;370(1677):20140212. doi: 10.1098/rstb.2014.0212. Philos Trans R Soc Lond B Biol Sci. 2015. PMID: 26240427 Free PMC article. Review.
-
Microbial Rhodopsin Optogenetic Tools: Application for Analyses of Synaptic Transmission and of Neuronal Network Activity in Behavior.Methods Mol Biol. 2015;1327:87-103. doi: 10.1007/978-1-4939-2842-2_8. Methods Mol Biol. 2015. PMID: 26423970
-
Sensory modulation of neuropeptide signaling by CASY-1 gates cholinergic transmission at Caenorhabditis elegans neuromuscular junction.J Biosci. 2025;50:4. doi: 10.1007/s12038-024-00488-x. J Biosci. 2025. PMID: 39912398 Free PMC article.
-
Japanese studies on neural circuits and behavior of Caenorhabditis elegans.Front Neural Circuits. 2013 Nov 28;7:187. doi: 10.3389/fncir.2013.00187. Front Neural Circuits. 2013. PMID: 24348340 Free PMC article. Review.
-
Optogenetic manipulation of neural activity in C. elegans: from synapse to circuits and behaviour.Biol Cell. 2013 Jun;105(6):235-50. doi: 10.1111/boc.201200069. Epub 2013 Apr 26. Biol Cell. 2013. PMID: 23458457 Review.
Cited by
-
Input density tunes Kenyon cell sensory responses in the Drosophila mushroom body.Curr Biol. 2023 Jul 10;33(13):2742-2760.e12. doi: 10.1016/j.cub.2023.05.064. Epub 2023 Jun 21. Curr Biol. 2023. PMID: 37348501 Free PMC article.
-
Bioluminescence as a functional tool for visualizing and controlling neuronal activity in vivo.Neurophotonics. 2024 Apr;11(2):024203. doi: 10.1117/1.NPh.11.2.024203. Epub 2024 Feb 12. Neurophotonics. 2024. PMID: 38348359 Free PMC article.
-
Hacking brain development to test models of sensory coding.bioRxiv [Preprint]. 2023 Jan 26:2023.01.25.525425. doi: 10.1101/2023.01.25.525425. bioRxiv. 2023. PMID: 36747712 Free PMC article. Preprint.
-
Understanding neural circuit function through synaptic engineering.Nat Rev Neurosci. 2024 Feb;25(2):131-139. doi: 10.1038/s41583-023-00777-8. Epub 2024 Jan 3. Nat Rev Neurosci. 2024. PMID: 38172626 Review.
-
Selective control of synaptically-connected circuit elements by all-optical synapses.Commun Biol. 2022 Jan 11;5(1):33. doi: 10.1038/s42003-021-02981-7. Commun Biol. 2022. PMID: 35017641 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
