

Quantitative imaging of RNA polymerase II activity in plants reveals the single-cell basis of tissue-wide transcriptional dynamics
- PMID: 34373604
- PMCID: PMC8616715
- DOI: 10.1038/s41477-021-00976-0
Quantitative imaging of RNA polymerase II activity in plants reveals the single-cell basis of tissue-wide transcriptional dynamics
Abstract
The responses of plants to their environment are often dependent on the spatiotemporal dynamics of transcriptional regulation. While live-imaging tools have been used extensively to quantitatively capture rapid transcriptional dynamics in living animal cells, the lack of implementation of these technologies in plants has limited concomitant quantitative studies in this kingdom. Here, we applied the PP7 and MS2 RNA-labelling technologies for the quantitative imaging of RNA polymerase II activity dynamics in single cells of living plants as they respond to experimental treatments. Using this technology, we counted nascent RNA transcripts in real time in Nicotiana benthamiana (tobacco) and Arabidopsis thaliana. Examination of heat shock reporters revealed that plant tissues respond to external signals by modulating the proportion of cells that switch from an undetectable basal state to a high-transcription state, instead of modulating the rate of transcription across all cells in a graded fashion. This switch-like behaviour, combined with cell-to-cell variability in transcription rate, results in mRNA production variability spanning three orders of magnitude. We determined that cellular heterogeneity stems mainly from stochasticity intrinsic to individual alleles instead of variability in cellular composition. Together, our results demonstrate that it is now possible to quantitatively study the dynamics of transcriptional programs in single cells of living plants.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
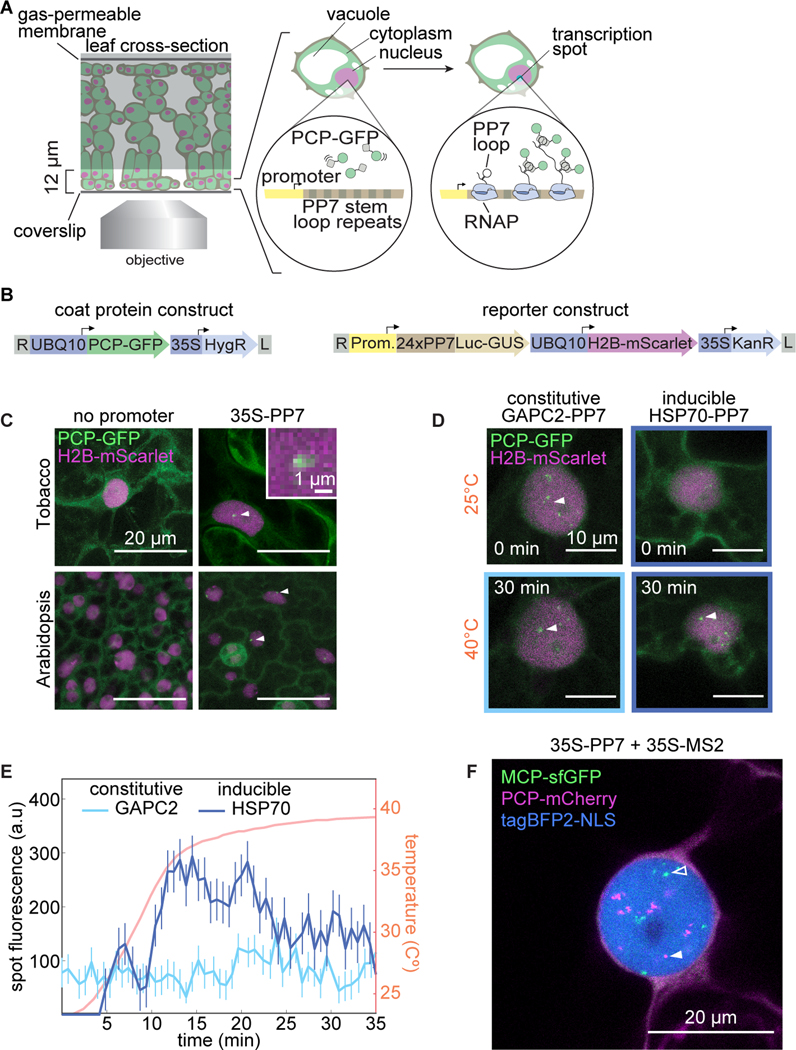
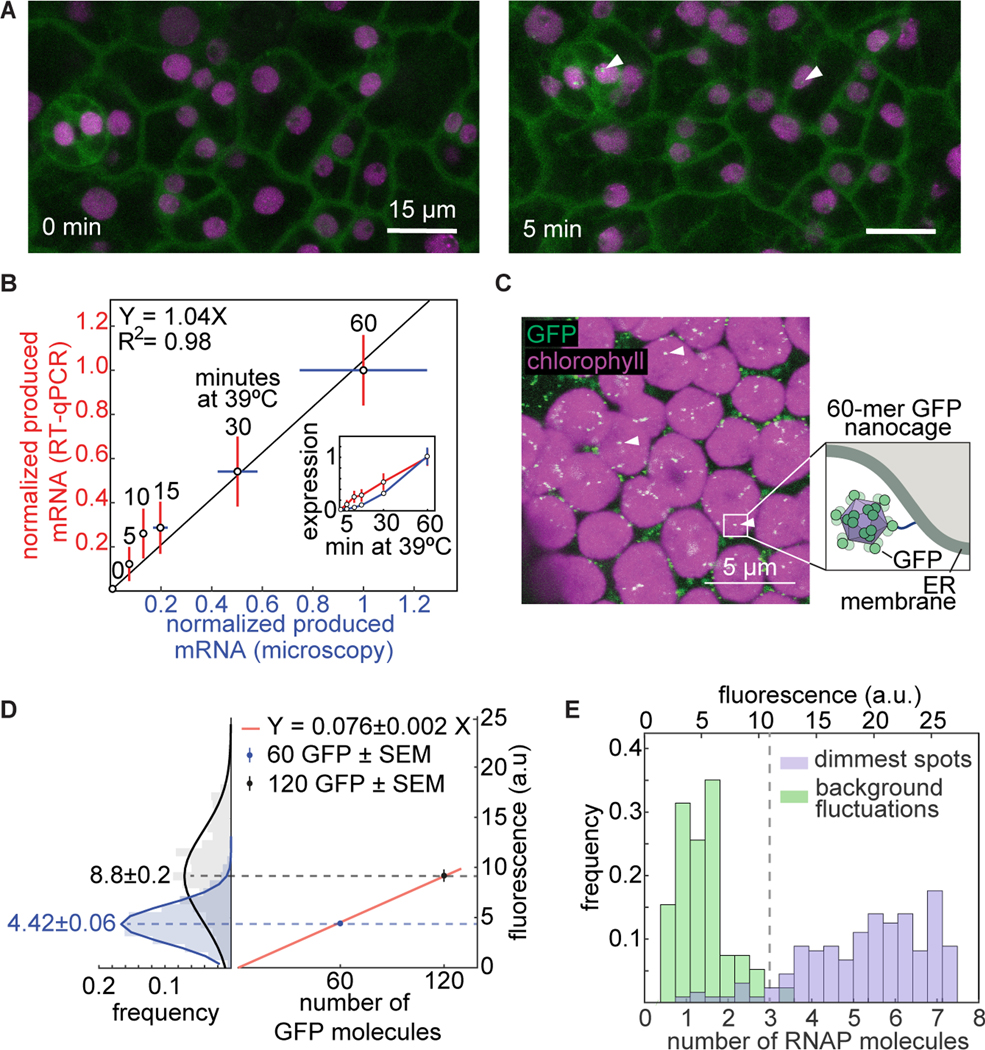
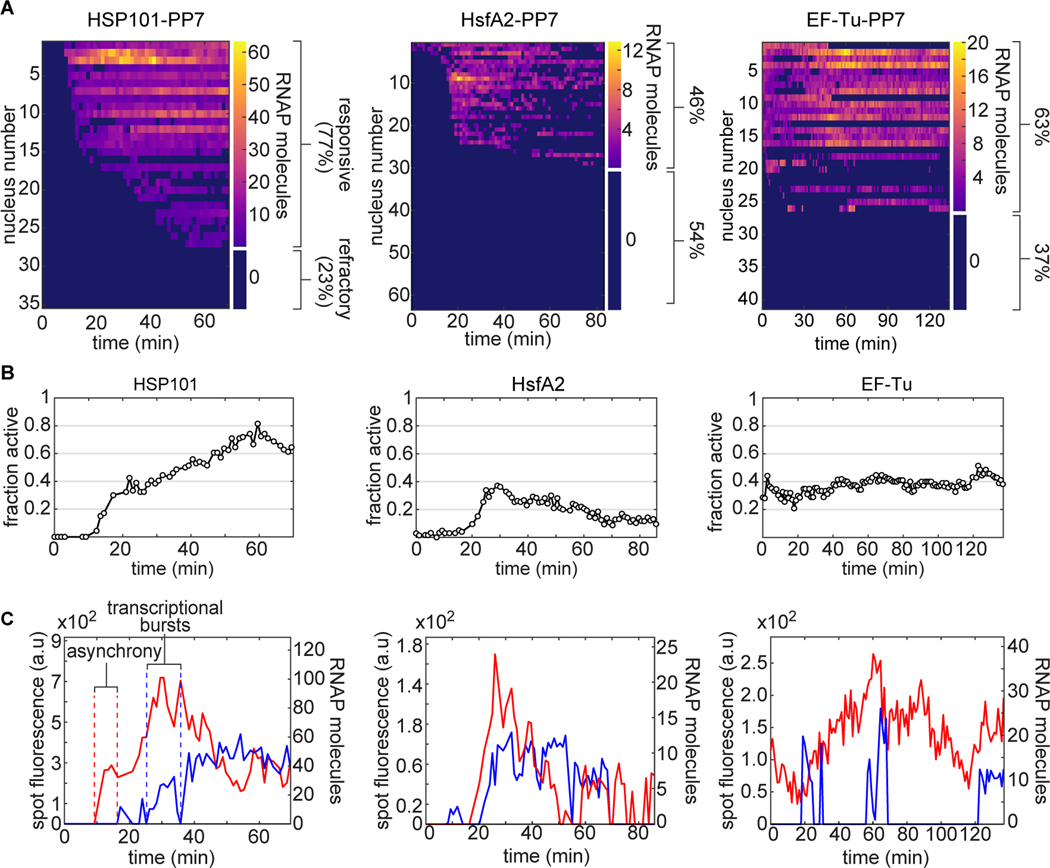

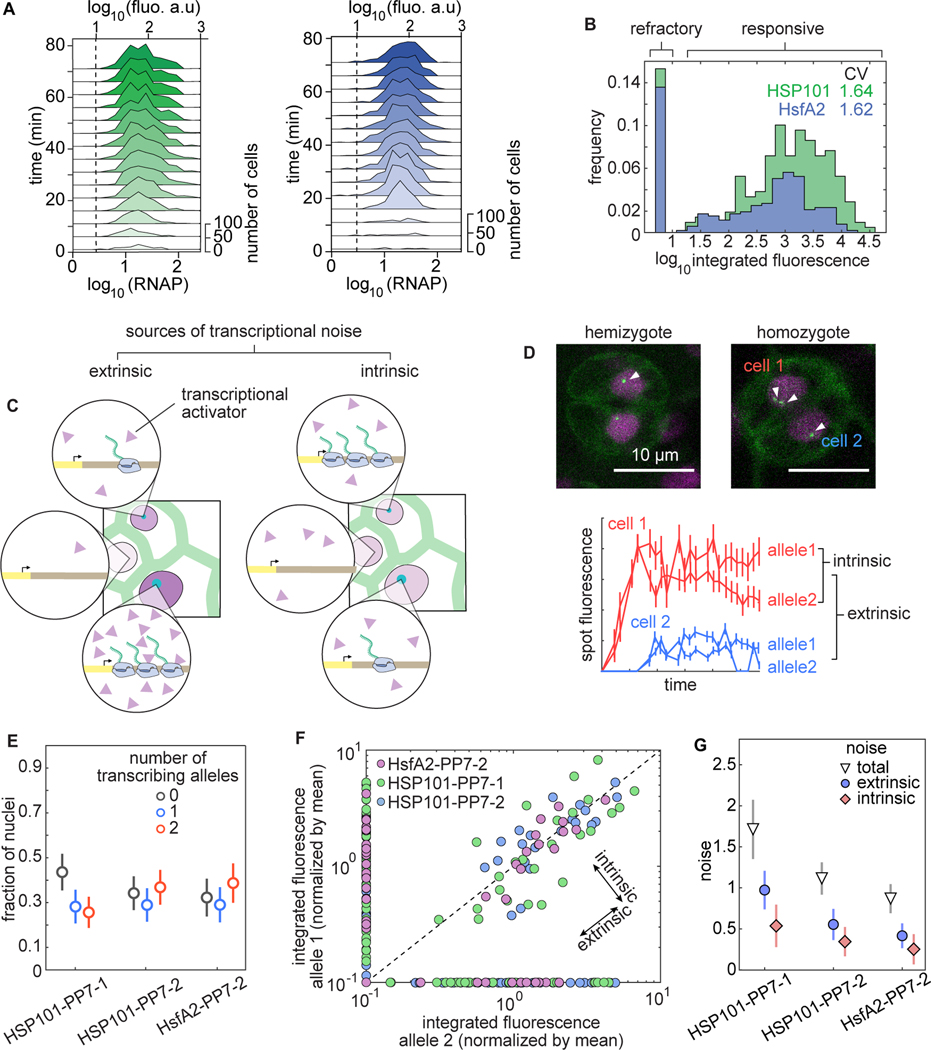
Comment in
-
Noisy transcription under the spotlight.Nat Plants. 2021 Aug;7(8):996-997. doi: 10.1038/s41477-021-00987-x. Nat Plants. 2021. PMID: 34373606 No abstract available.
References
-
- Angel A, Song J, Dean C, and Howard M. (2011). A Polycomb-based switch underlying quantitative epigenetic memory. Nature, 476(7358):105–109. - PubMed
-
- Ardehali MB and Lis JT (2009). Tracking rates of transcription and splicing in vivo. Nature Structural and Molecular Biology, 16(11):1123–1124. - PubMed
-
- Arganda-Carreras I, Kaynig V, Rueden C, Eliceiri KW, Schindelin J, Cardona A, and Seung HS (2017). Trainable Weka Segmentation: A machine learning tool for microscopy pixel classification. Bioinformatics, 33(15):2424–2426. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
