

Mucus
- PMID: 34375594
- PMCID: PMC8759706
- DOI: 10.1016/j.cub.2021.06.093
Mucus
Abstract
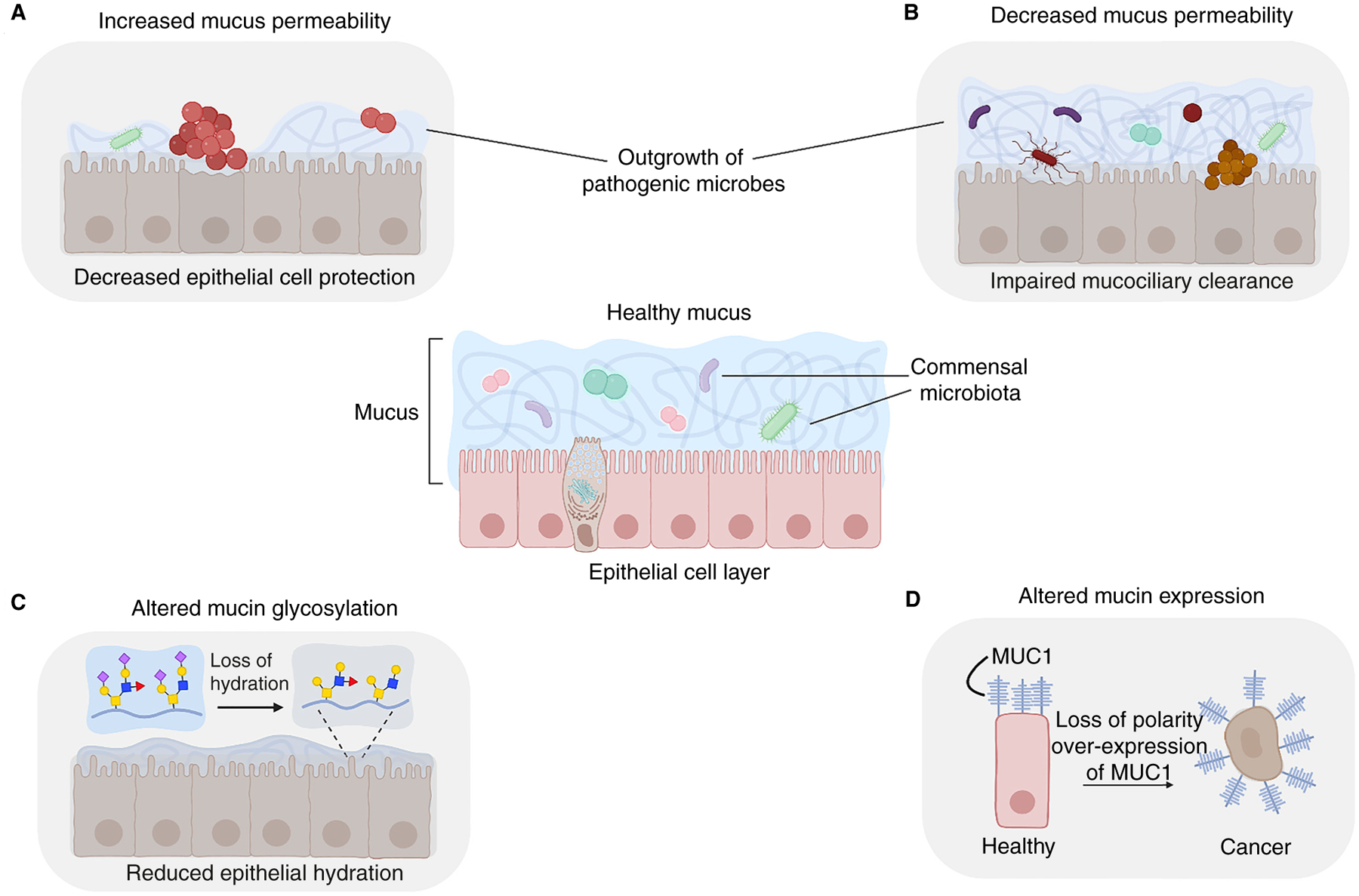
Mucus is a slimy hydrogel that lines the mucosal surfaces in our body, including the intestines, stomach, eyes, lungs and urogenital tract. This glycoprotein-rich network is truly the jack of all trades. As a barrier, it lubricates surfaces, protects our cells from physical stress, and selectively allows the passage of nutrients while clearing out pathogens and debris. As a home to our microbiota, it supports a level of microbial diversity that is unattainable with most culture methods. As a reservoir of complex carbohydrate structures called glycans, it plays critical roles in controlling cell adhesion and signaling, and it alters the behavior and spatial distribution of microbes. On top of all this, mucus regulates the passage of sperm during fertilization, heals wounds, helps us smell, and prevents the stomach from digesting itself, to name just a few of its functions. Given these impressive features, it is no wonder that mucus crosses boundaries of species and kingdoms - mucus gels are made by organisms ranging from the simplest metazoans to corals, snails, fish, and frogs. It is also no surprise that mucus is exploited in everyday applications, including foods, cosmetics, and other products relevant to medicine and industry.
Copyright © 2021. Published by Elsevier Inc.
Figures
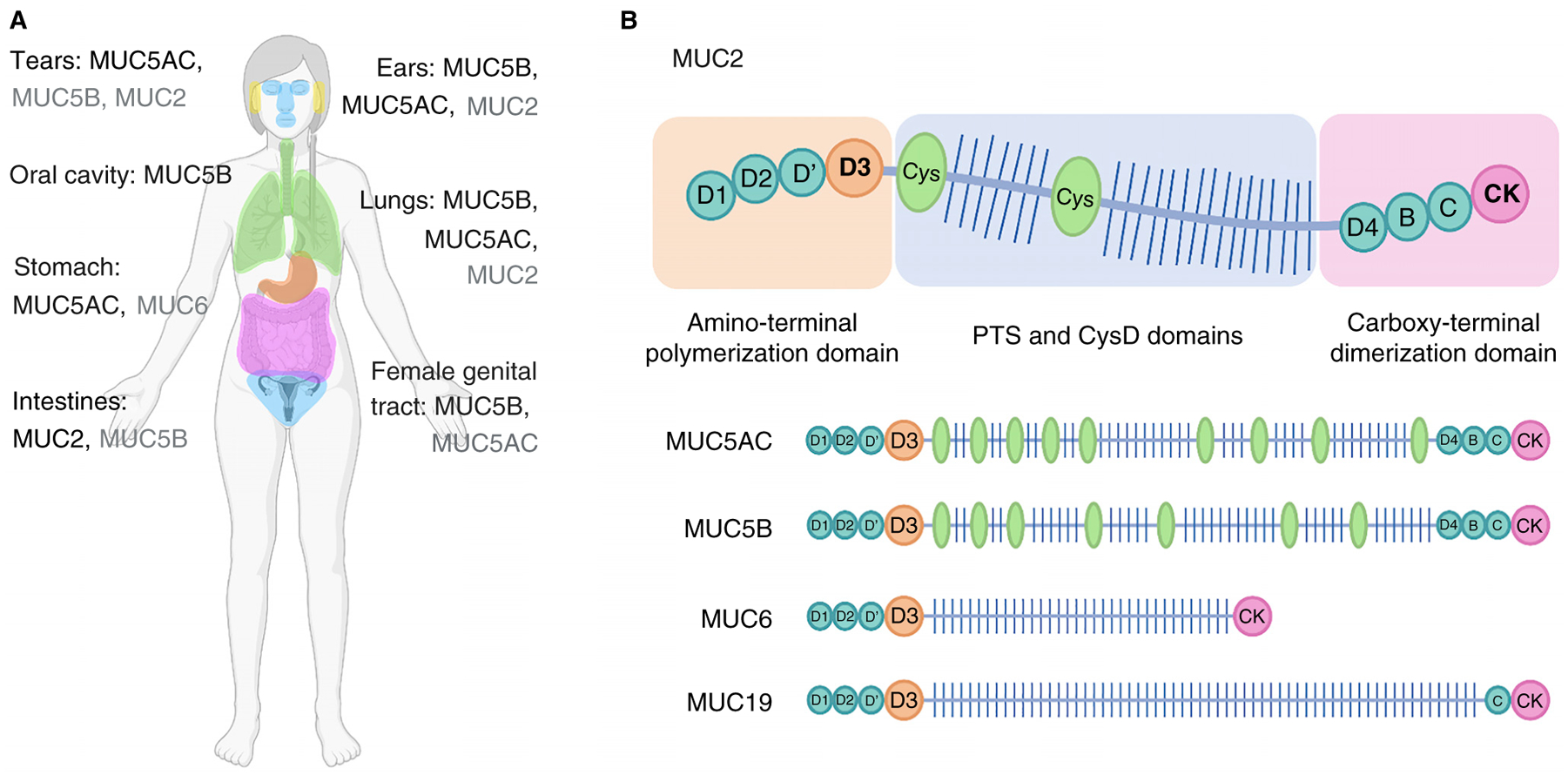
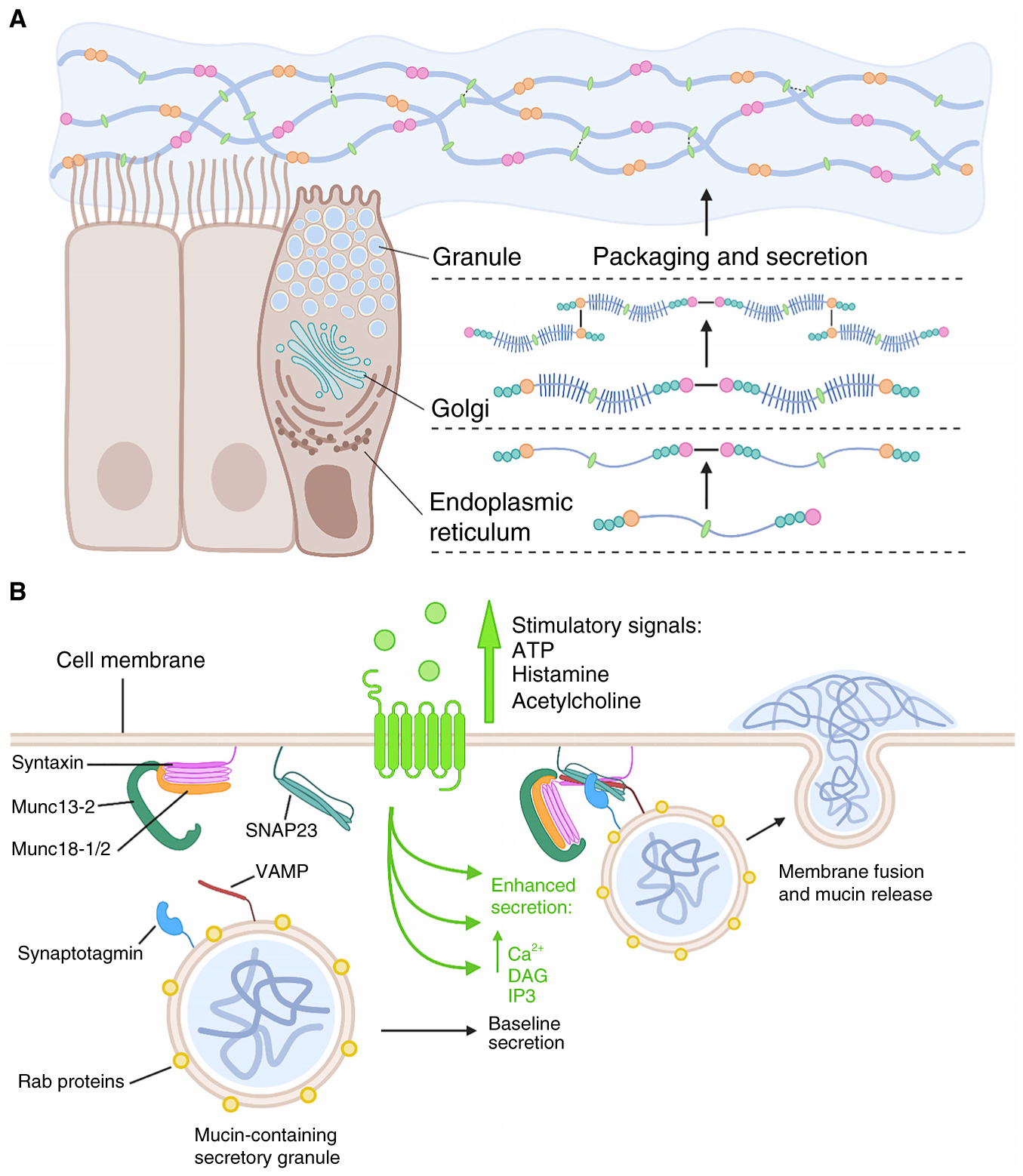
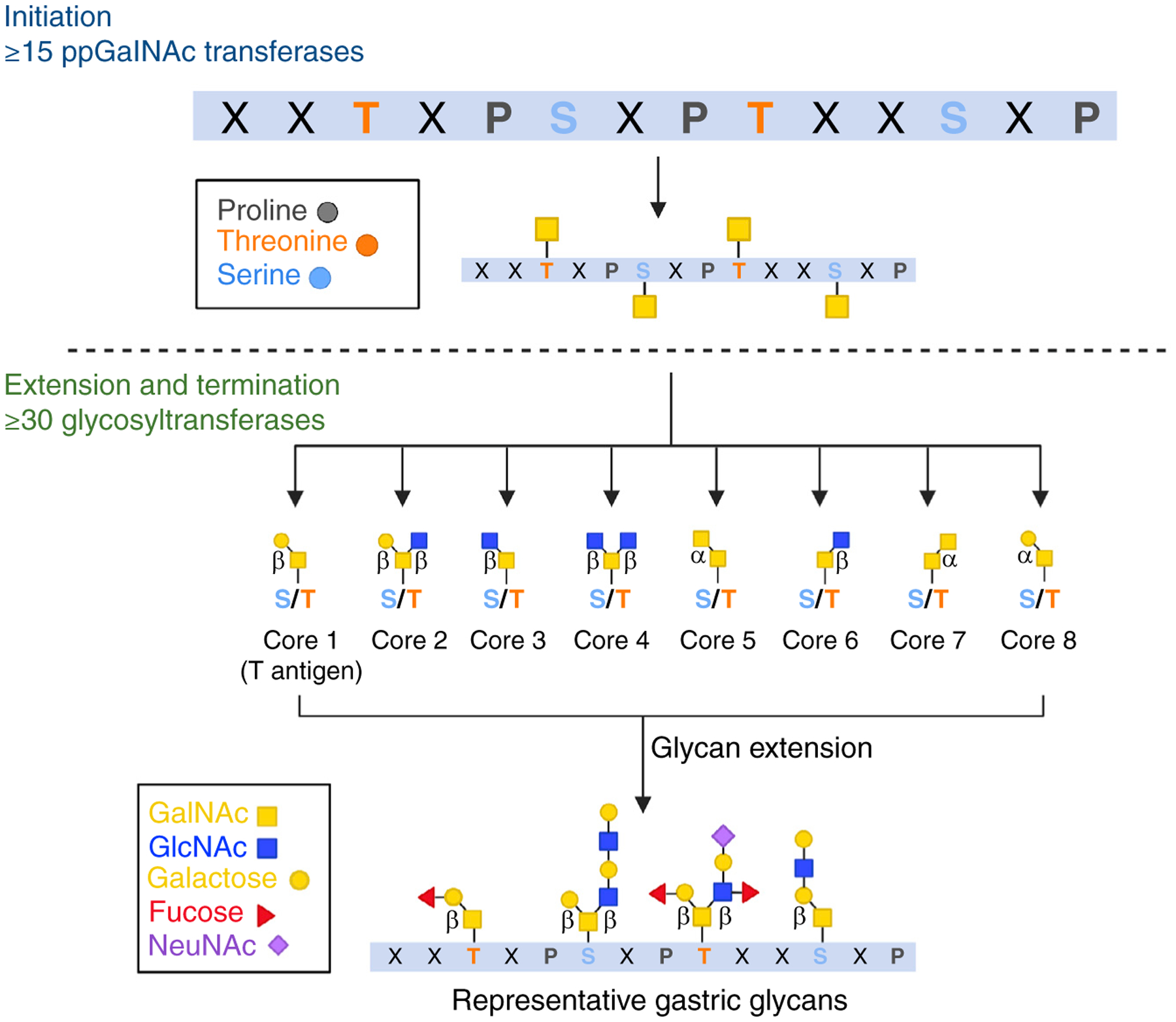
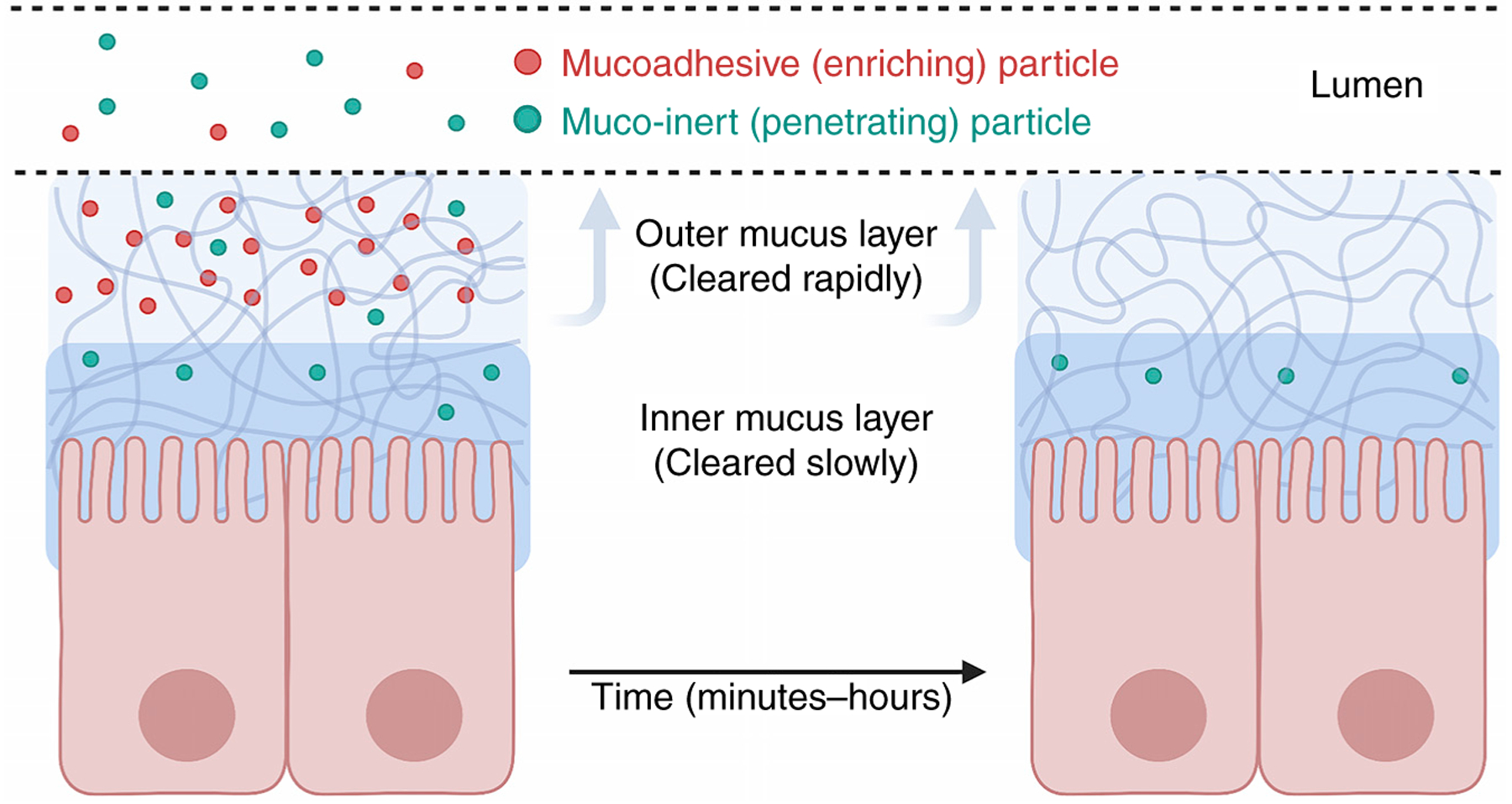
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
