

Mapping of pseudouridine residues on cellular and viral transcripts using a novel antibody-based technique
- PMID: 34376564
- PMCID: PMC8522693
- DOI: 10.1261/rna.078940.121
Mapping of pseudouridine residues on cellular and viral transcripts using a novel antibody-based technique
Abstract
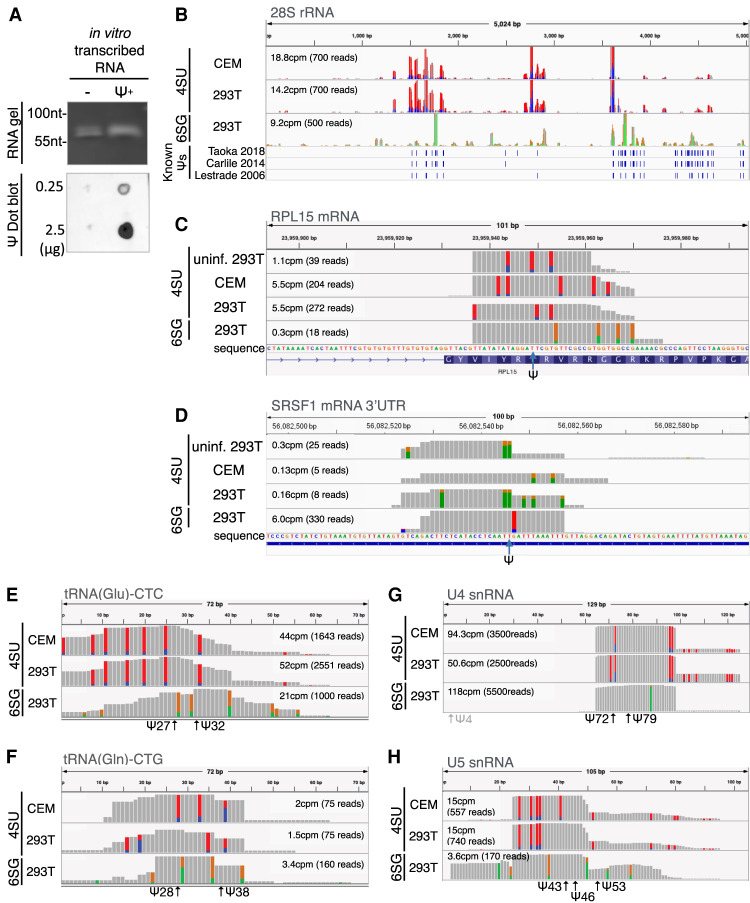
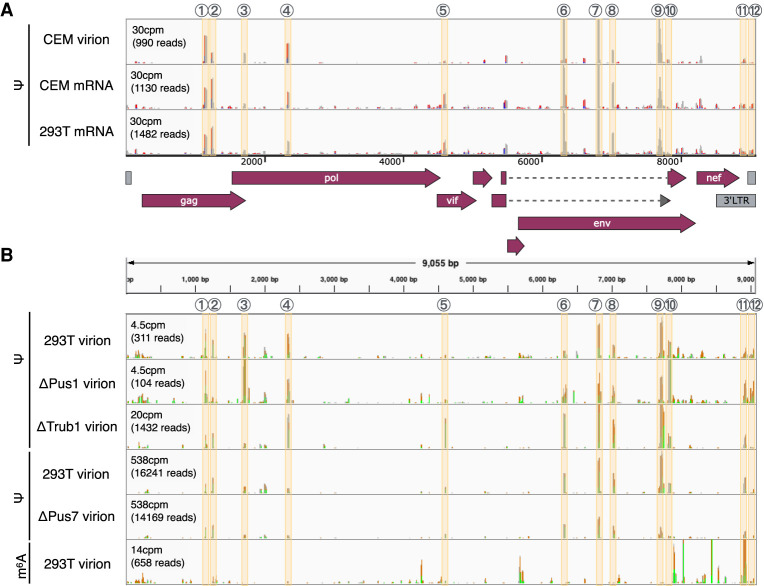
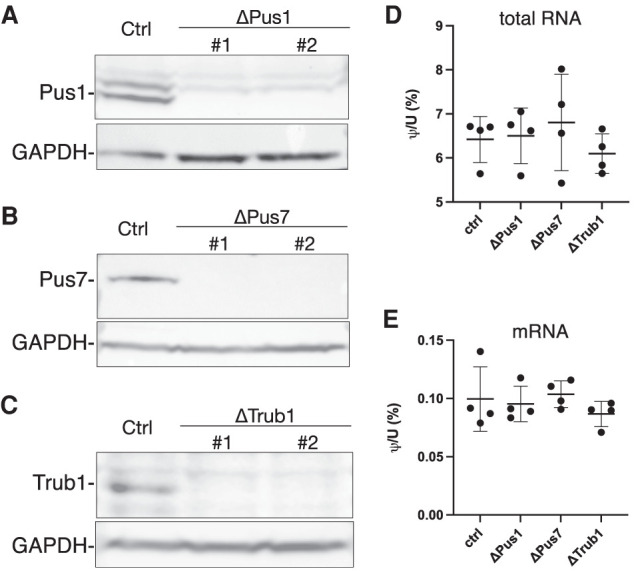
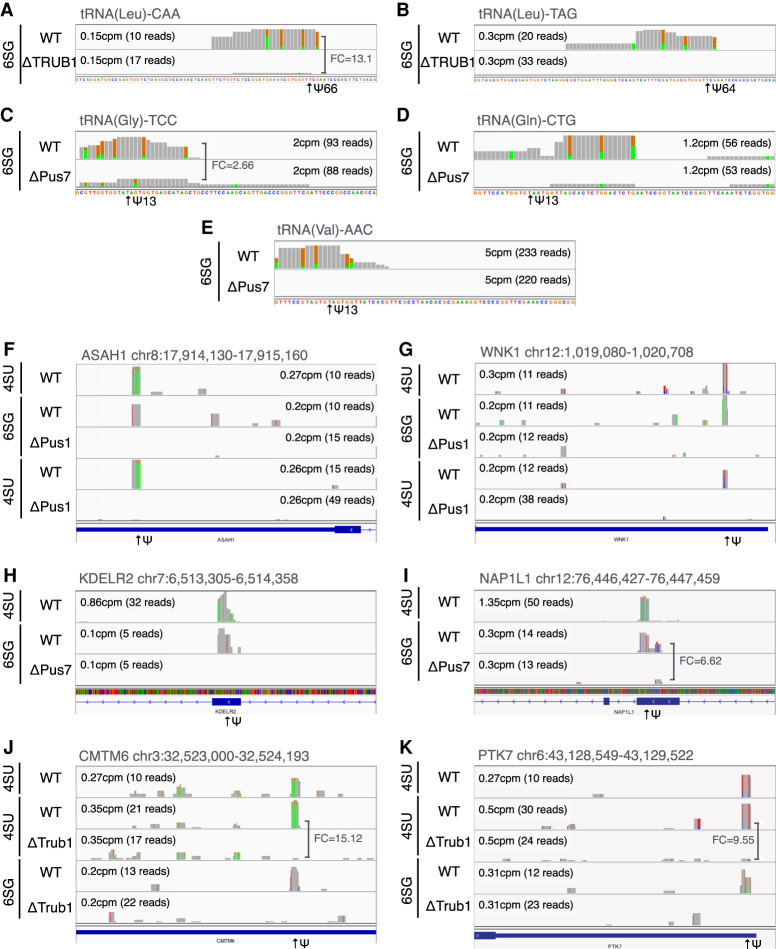
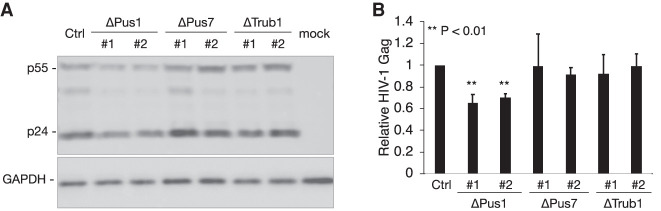
Pseudouridine (Ψ) is the most common noncanonical ribonucleoside present on mammalian noncoding RNAs (ncRNAs), including rRNAs, tRNAs, and snRNAs, where it contributes ∼7% of the total uridine level. However, Ψ constitutes only ∼0.1% of the uridines present on mRNAs and its effect on mRNA function remains unclear. Ψ residues have been shown to inhibit the detection of exogenous RNA transcripts by host innate immune factors, thus raising the possibility that viruses might have subverted the addition of Ψ residues to mRNAs by host pseudouridine synthase (PUS) enzymes as a way to inhibit antiviral responses in infected cells. Here, we describe and validate a novel antibody-based Ψ mapping technique called photo-crosslinking-assisted Ψ sequencing (PA-Ψ-seq) and use it to map Ψ residues on not only multiple cellular RNAs but also on the mRNAs and genomic RNA encoded by HIV-1. We describe 293T-derived cell lines in which human PUS enzymes previously reported to add Ψ residues to human mRNAs, specifically PUS1, PUS7, and TRUB1/PUS4, were inactivated by gene editing. Surprisingly, while this allowed us to assign several sites of Ψ addition on cellular mRNAs to each of these three PUS enzymes, Ψ sites present on HIV-1 transcripts remained unaffected. Moreover, loss of PUS1, PUS7, or TRUB1 function did not significantly reduce the level of Ψ residues detected on total human mRNA below the ∼0.1% level seen in wild-type cells, thus implying that the PUS enzyme(s) that adds the bulk of Ψ residues to human mRNAs remains to be defined.
Keywords: HIV-1; epitranscriptomics; post-transcriptional gene regulation; pseudouridine.
© 2021 Martinez Campos et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
Similar articles
-
Transcriptome-wide mapping of pseudouridines: pseudouridine synthases modify specific mRNAs in S. cerevisiae.PLoS One. 2014 Oct 29;9(10):e110799. doi: 10.1371/journal.pone.0110799. eCollection 2014. PLoS One. 2014. PMID: 25353621 Free PMC article.
-
Pseudouridines on Trypanosoma brucei mRNAs are developmentally regulated: Implications to mRNA stability and protein binding.Mol Microbiol. 2021 Sep;116(3):808-826. doi: 10.1111/mmi.14774. Epub 2021 Jul 7. Mol Microbiol. 2021. PMID: 34165831
-
Identifying Individual Pseudouridine (Ψ) Sites Across Transcripts from HIV-1 Infected Cells.Methods Mol Biol. 2024;2807:229-242. doi: 10.1007/978-1-0716-3862-0_16. Methods Mol Biol. 2024. PMID: 38743232
-
Research progress of RNA pseudouridine modification in nervous system.Int J Neurosci. 2025 Jun;135(6):639-649. doi: 10.1080/00207454.2024.2315483. Epub 2024 Feb 29. Int J Neurosci. 2025. PMID: 38407188 Review.
-
Pseudouridine: still mysterious, but never a fake (uridine)!RNA Biol. 2014;11(12):1540-54. doi: 10.4161/15476286.2014.992278. RNA Biol. 2014. PMID: 25616362 Free PMC article. Review.
Cited by
-
Ribosomal RNA Pseudouridylation: Will Newly Available Methods Finally Define the Contribution of This Modification to Human Ribosome Plasticity?Front Genet. 2022 Jun 1;13:920987. doi: 10.3389/fgene.2022.920987. eCollection 2022. Front Genet. 2022. PMID: 35719370 Free PMC article. Review.
-
Past, Present, and Future of RNA Modifications in Infectious Disease Research.ACS Infect Dis. 2024 Dec 13;10(12):4017-4029. doi: 10.1021/acsinfecdis.4c00598. Epub 2024 Nov 21. ACS Infect Dis. 2024. PMID: 39569943
-
Epitranscriptomic cytidine methylation of the hepatitis B viral RNA is essential for viral reverse transcription and particle production.Proc Natl Acad Sci U S A. 2024 Jun 11;121(24):e2400378121. doi: 10.1073/pnas.2400378121. Epub 2024 Jun 3. Proc Natl Acad Sci U S A. 2024. PMID: 38830096 Free PMC article.
-
Control of protein synthesis through mRNA pseudouridylation by dyskerin.Sci Adv. 2023 Jul 28;9(30):eadg1805. doi: 10.1126/sciadv.adg1805. Epub 2023 Jul 28. Sci Adv. 2023. PMID: 37506213 Free PMC article.
-
Nanopore Dwell Time Analysis Permits Sequencing and Conformational Assignment of Pseudouridine in SARS-CoV-2.ACS Cent Sci. 2021 Oct 27;7(10):1707-1717. doi: 10.1021/acscentsci.1c00788. Epub 2021 Sep 15. ACS Cent Sci. 2021. PMID: 34729414 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases