

A Methyltransferase-Defective Vesicular Stomatitis Virus-Based SARS-CoV-2 Vaccine Candidate Provides Complete Protection against SARS-CoV-2 Infection in Hamsters
- PMID: 34379509
- PMCID: PMC8475528
- DOI: 10.1128/JVI.00592-21
A Methyltransferase-Defective Vesicular Stomatitis Virus-Based SARS-CoV-2 Vaccine Candidate Provides Complete Protection against SARS-CoV-2 Infection in Hamsters
Abstract
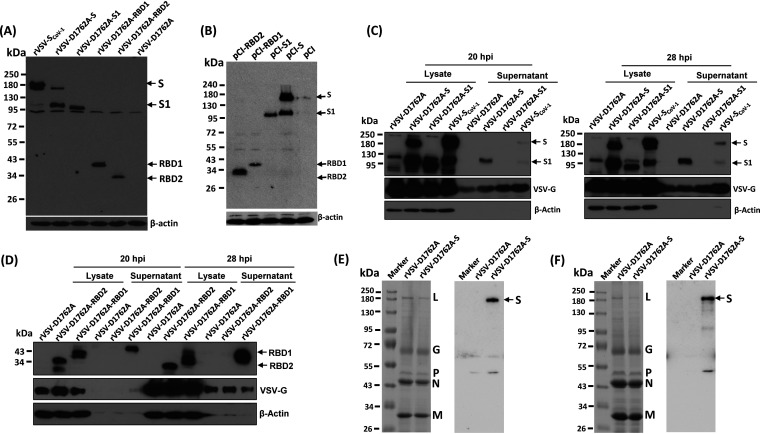
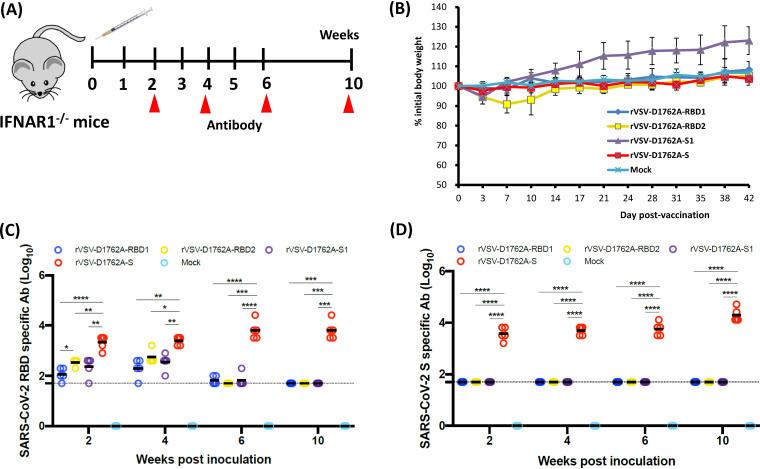
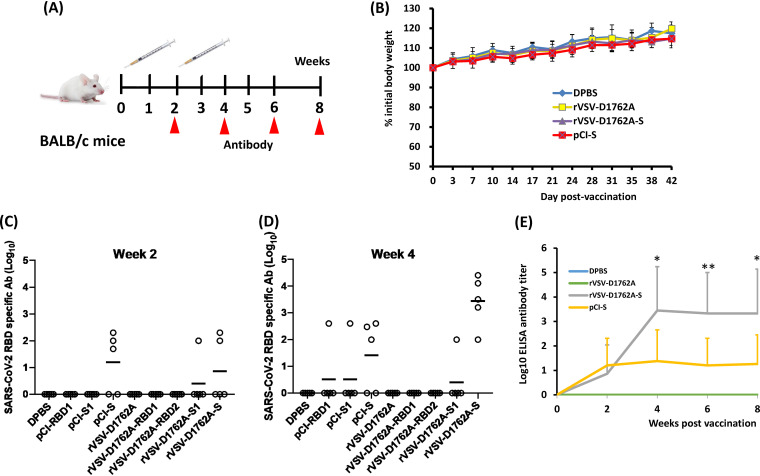
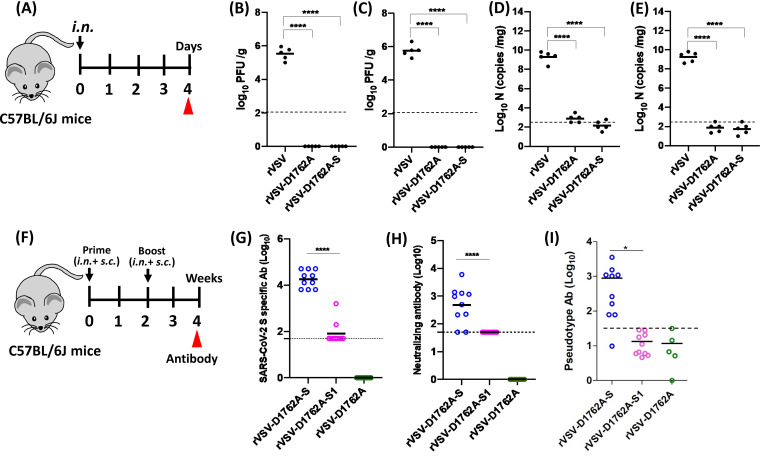
The current pandemic of coronavirus disease 2019 (COVID-19) caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has led to dramatic economic and health burdens. Although the worldwide SARS-CoV-2 vaccination campaign has begun, exploration of other vaccine candidates is needed due to uncertainties with the current approved vaccines, such as durability of protection, cross-protection against variant strains, and costs of long-term production and storage. In this study, we developed a methyltransferase-defective recombinant vesicular stomatitis virus (mtdVSV)-based SARS-CoV-2 vaccine candidate. We generated mtdVSVs expressing SARS-CoV-2 full-length spike (S) protein, S1, or its receptor-binding domain (RBD). All of these recombinant viruses grew to high titers in mammalian cells despite high attenuation in cell culture. The SARS-CoV-2 S protein and its truncations were highly expressed by the mtdVSV vector. These mtdVSV-based vaccine candidates were completely attenuated in both immunocompetent and immunocompromised mice. Among these constructs, mtdVSV-S induced high levels of SARS-CoV-2-specific neutralizing antibodies (NAbs) and Th1-biased T-cell immune responses in mice. In Syrian golden hamsters, the serum levels of SARS-CoV-2-specific NAbs triggered by mtdVSV-S were higher than the levels of NAbs in convalescent plasma from recovered COVID-19 patients. In addition, hamsters immunized with mtdVSV-S were completely protected against SARS-CoV-2 replication in lung and nasal turbinate tissues, cytokine storm, and lung pathology. Collectively, our data demonstrate that mtdVSV expressing SARS-CoV-2 S protein is a safe and highly efficacious vaccine candidate against SARS-CoV-2 infection. IMPORTANCE Viral mRNA cap methyltransferase (MTase) is essential for mRNA stability, protein translation, and innate immune evasion. Thus, viral mRNA cap MTase activity is an excellent target for development of live attenuated or live vectored vaccine candidates. Here, we developed a panel of MTase-defective recombinant vesicular stomatitis virus (mtdVSV)-based SARS-CoV-2 vaccine candidates expressing full-length S, S1, or several versions of the RBD. These mtdVSV-based vaccine candidates grew to high titers in cell culture and were completely attenuated in both immunocompetent and immunocompromised mice. Among these vaccine candidates, mtdVSV-S induces high levels of SARS-CoV-2-specific neutralizing antibodies (Nabs) and Th1-biased immune responses in mice. Syrian golden hamsters immunized with mtdVSV-S triggered SARS-CoV-2-specific NAbs at higher levels than those in convalescent plasma from recovered COVID-19 patients. Furthermore, hamsters immunized with mtdVSV-S were completely protected against SARS-CoV-2 challenge. Thus, mtdVSV is a safe and highly effective vector to deliver SARS-CoV-2 vaccine.
Keywords: SARS-CoV-2; VSV; mRNA cap methyltransferase; vaccine.
Figures







Similar articles
-
Immunogenicity and efficacy of XBB.1.5 rS vaccine against the EG.5.1 variant of SARS-CoV-2 in Syrian hamsters.J Virol. 2024 Oct 22;98(10):e0052824. doi: 10.1128/jvi.00528-24. Epub 2024 Sep 4. J Virol. 2024. PMID: 39230305 Free PMC article.
-
A safe and highly efficacious measles virus-based vaccine expressing SARS-CoV-2 stabilized prefusion spike.Proc Natl Acad Sci U S A. 2021 Mar 23;118(12):e2026153118. doi: 10.1073/pnas.2026153118. Proc Natl Acad Sci U S A. 2021. PMID: 33688034 Free PMC article.
-
Mucosal SARS-CoV-2 S1 adenovirus-based vaccine elicits robust systemic and mucosal immunity and protects against disease in animals.mBio. 2025 Jan 8;16(1):e0217024. doi: 10.1128/mbio.02170-24. Epub 2024 Dec 4. mBio. 2025. PMID: 39629990 Free PMC article.
-
Severe acute respiratory syndrome-coronavirus-2 spike (S) protein based vaccine candidates: State of the art and future prospects.Rev Med Virol. 2021 May;31(3):e2183. doi: 10.1002/rmv.2183. Epub 2020 Oct 15. Rev Med Virol. 2021. PMID: 33594794 Free PMC article. Review.
-
Emerging severe acute respiratory syndrome coronavirus 2 variants and their impact on immune evasion and vaccine-induced immunity.Trans R Soc Trop Med Hyg. 2024 Dec 3;118(12):761-772. doi: 10.1093/trstmh/trae060. Trans R Soc Trop Med Hyg. 2024. PMID: 39297227 Review.
Cited by
-
Fighting Fire with Fire: Immunogenicity of Viral Vectored Vaccines against COVID-19.Viruses. 2022 Feb 12;14(2):380. doi: 10.3390/v14020380. Viruses. 2022. PMID: 35215973 Free PMC article. Review.
-
Pseudotyped Vesicular Stomatitis Virus-Severe Acute Respiratory Syndrome-Coronavirus-2 Spike for the Study of Variants, Vaccines, and Therapeutics Against Coronavirus Disease 2019.Front Microbiol. 2022 Jan 14;12:817200. doi: 10.3389/fmicb.2021.817200. eCollection 2021. Front Microbiol. 2022. PMID: 35095820 Free PMC article. Review.
-
Viral vectored vaccines: design, development, preventive and therapeutic applications in human diseases.Signal Transduct Target Ther. 2023 Apr 7;8(1):149. doi: 10.1038/s41392-023-01408-5. Signal Transduct Target Ther. 2023. PMID: 37029123 Free PMC article. Review.
-
Viral Vector Vaccine Development and Application during the COVID-19 Pandemic.Microorganisms. 2022 Jul 18;10(7):1450. doi: 10.3390/microorganisms10071450. Microorganisms. 2022. PMID: 35889169 Free PMC article. Review.
-
Three SARS-CoV-2 spike protein variants delivered intranasally by measles and mumps vaccines are broadly protective.Nat Commun. 2024 Jul 3;15(1):5589. doi: 10.1038/s41467-024-49443-2. Nat Commun. 2024. PMID: 38961063 Free PMC article.
References
-
- Li Q, Guan X, Wu P, Wang X, Zhou L, Tong Y, Ren R, Leung KSM, Lau EHY, Wong JY, Xing X, Xiang N, Wu Y, Li C, Chen Q, Li D, Liu T, Zhao J, Liu M, Tu W, Chen C, Jin L, Yang R, Wang Q, Zhou S, Wang R, Liu H, Luo Y, Liu Y, Shao G, Li H, Tao Z, Yang Y, Deng Z, Liu B, Ma Z, Zhang Y, Shi G, Lam TTY, Wu JT, Gao GF, Cowling BJ, Yang B, Leung GM, Feng Z. 2020. Early transmission dynamics in Wuhan, China, of novel coronavirus-infected pneumonia. N Engl J Med 382:1199–1207. 10.1056/NEJMoa2001316. - DOI - PMC - PubMed
-
- Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, Zhang L, Fan G, Xu J, Gu X, Cheng Z, Yu T, Xia J, Wei Y, Wu W, Xie X, Yin W, Li H, Liu M, Xiao Y, Gao H, Guo L, Xie J, Wang G, Jiang R, Gao Z, Jin Q, Wang J, Cao B. 2020. Clinical features of patients infected with 2019 novel coronavirus in Wuhan. Lancet 395:497–506. 10.1016/S0140-6736(20)30183-5. - DOI - PMC - PubMed
-
- Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, Zhao X, Huang B, Shi W, Lu R, Niu P, Zhan F, Ma X, Wang D, Xu W, Wu G, Gao GF, Tan W, China Novel Coronavirus Investigating and Research Team. 2020. A Novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med 382:727–733. 10.1056/NEJMoa2001017. - DOI - PMC - PubMed
-
- Kemp SA, Collier DA, Datir RP, Ferreira IATM, Gayed S, Jahun A, Hosmillo M, Rees-Spear C, Mlcochova P, Lumb IU, Roberts DJ, Chandra A, Temperton N, Sharrocks K, Blane E, Modis Y, Leigh KE, Briggs JAG, van Gils MJ, Smith KGC, Bradley JR, Smith C, Doffinger R, Ceron-Gutierrez L, Barcenas-Morales G, Pollock DD, Goldstein RA, Smielewska A, Skittrall JP, Gouliouris T, Goodfellow IG, Gkrania-Klotsas E, Illingworth CJR, McCoy LE, Gupta RK, COVID-19 Genomics UK (COG-UK) Consortium. 2021. SARS-CoV-2 evolution during treatment of chronic infection. Nature 592:277–282. 10.1038/s41586-021-03291-y. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI090060/AI/NIAID NIH HHS/United States
- R01 AI151175/AI/NIAID NIH HHS/United States
- R01 AI123661/AI/NIAID NIH HHS/United States
- P01 AI112524/AI/NIAID NIH HHS/United States
- R01 AI145144/AI/NIAID NIH HHS/United States
- R01 AI070723/AI/NIAID NIH HHS/United States
- R56 AI090060/AI/NIAID NIH HHS/United States
- R01 AI150473/AI/NIAID NIH HHS/United States
- R01 AI137567/AI/NIAID NIH HHS/United States
- R01 AI161175/AI/NIAID NIH HHS/United States
- R01 AI112381/AI/NIAID NIH HHS/United States
- RM1 HG008935/HG/NHGRI NIH HHS/United States
- U19 AI142733/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
