

Actinin BioID reveals sarcomere crosstalk with oxidative metabolism through interactions with IGF2BP2
- PMID: 34380038
- PMCID: PMC8447243
- DOI: 10.1016/j.celrep.2021.109512
Actinin BioID reveals sarcomere crosstalk with oxidative metabolism through interactions with IGF2BP2
Abstract
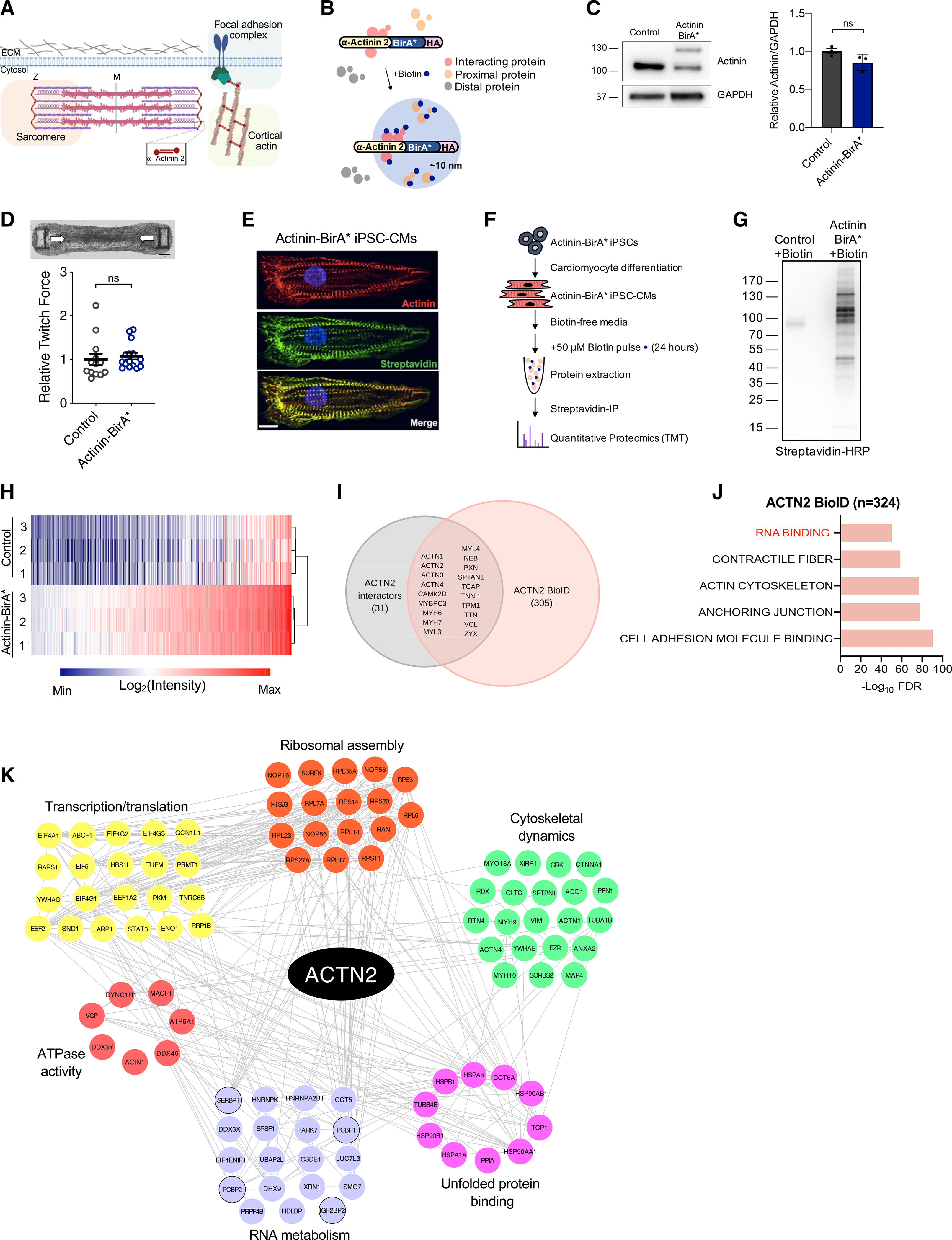
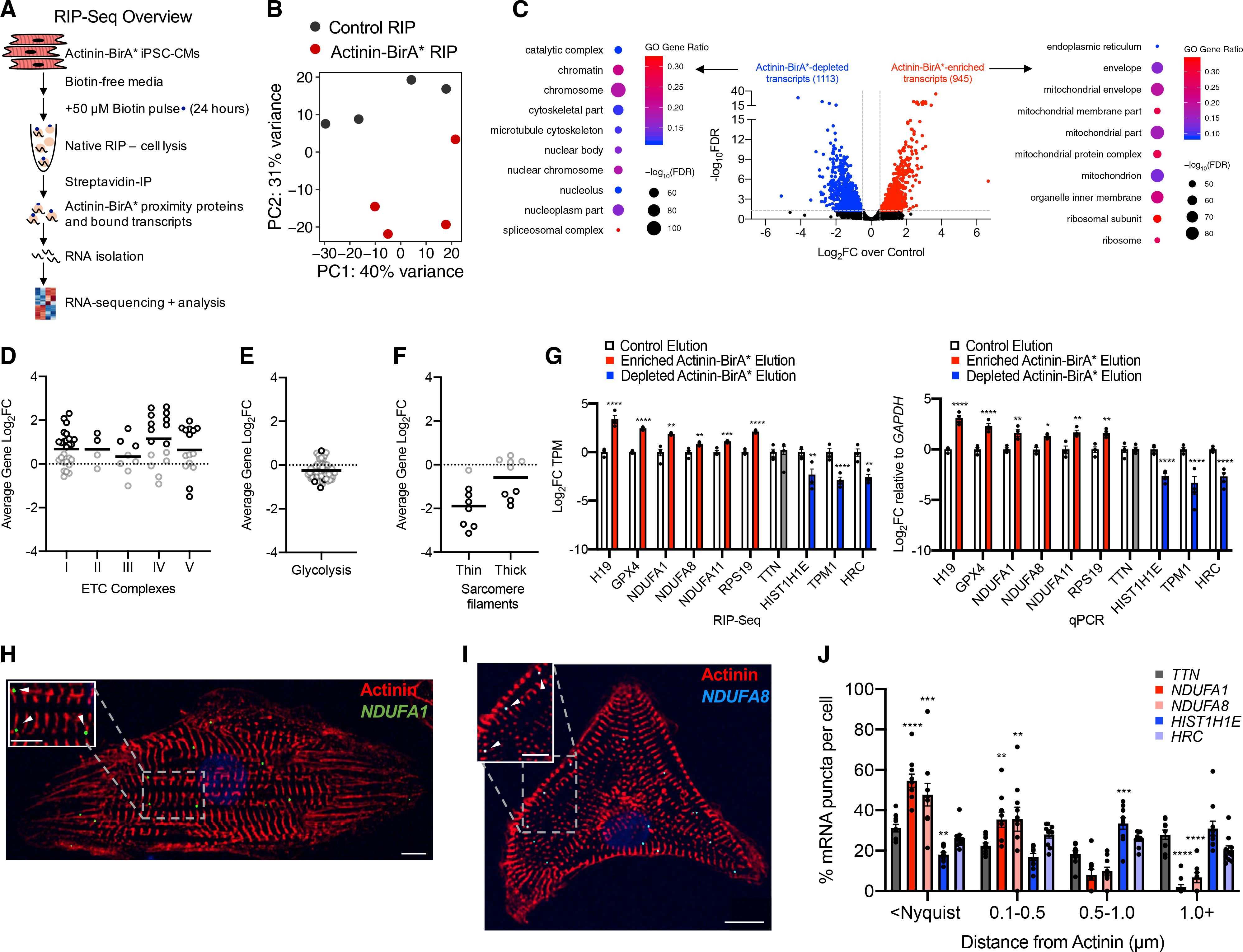
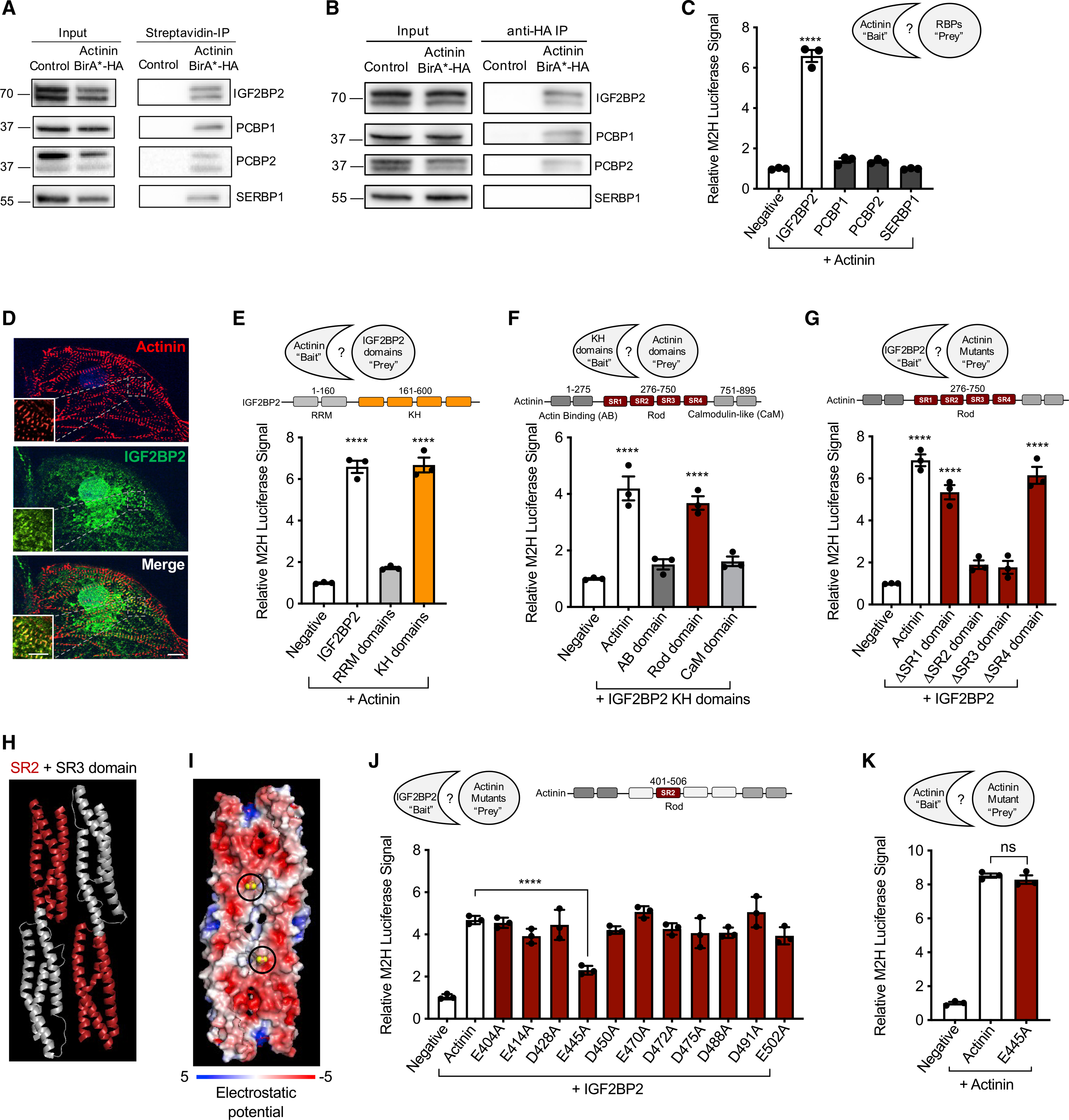
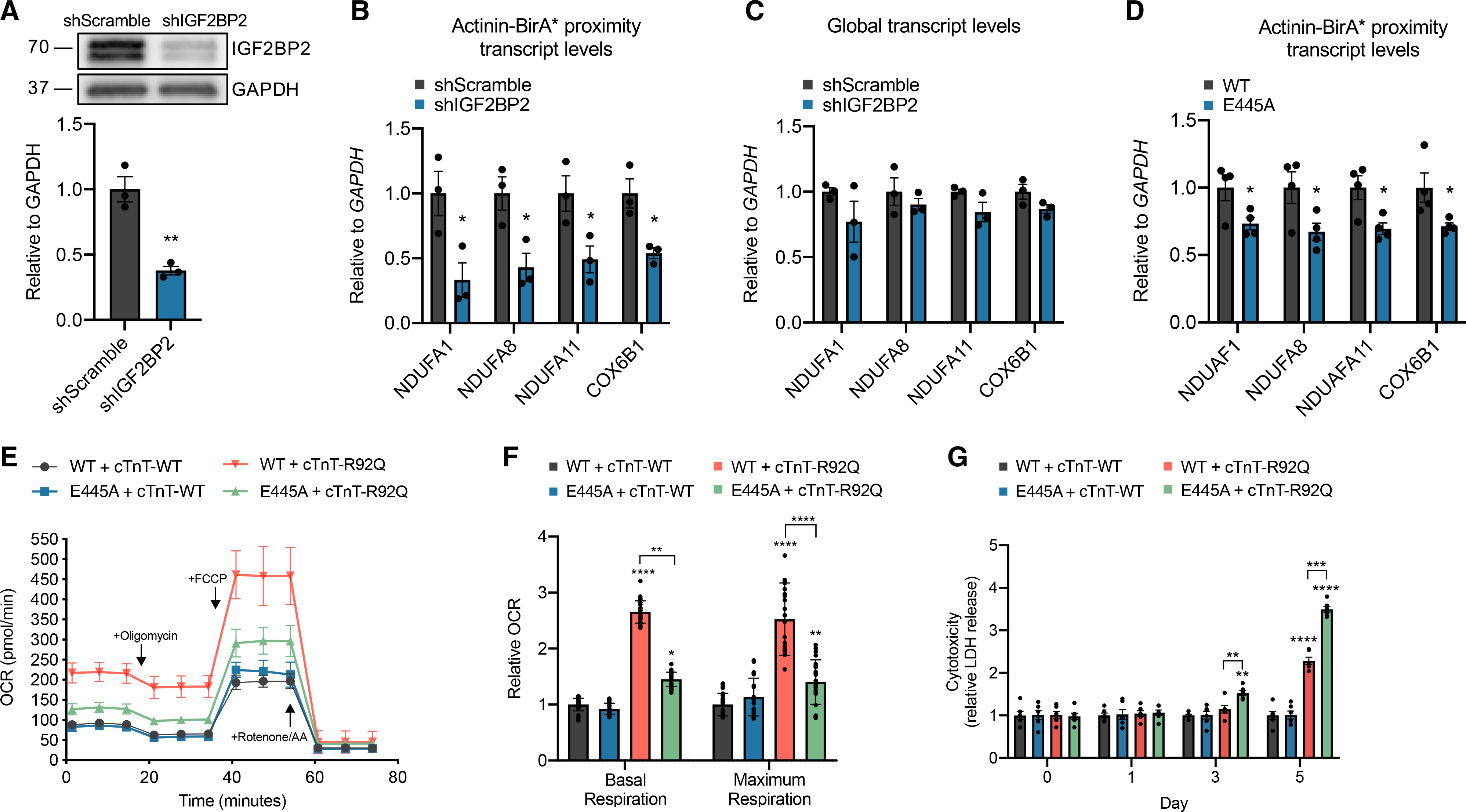
Actinins are strain-sensing actin cross-linkers that are ubiquitously expressed and harbor mutations in human diseases. We utilize CRISPR, pluripotent stem cells, and BioID to study actinin interactomes in human cardiomyocytes. We identify 324 actinin proximity partners, including those that are dependent on sarcomere assembly. We confirm 19 known interactors and identify a network of RNA-binding proteins, including those with RNA localization functions. In vivo and biochemical interaction studies support that IGF2BP2 localizes electron transport chain transcripts to actinin neighborhoods through interactions between its K homology (KH) domain and actinin's rod domain. We combine alanine scanning mutagenesis and metabolic assays to disrupt and functionally interrogate actinin-IGF2BP2 interactions, which reveal an essential role in metabolic responses to pathological sarcomere activation using a hypertrophic cardiomyopathy model. This study expands our functional knowledge of actinin, uncovers sarcomere interaction partners, and reveals sarcomere crosstalk with IGF2BP2 for metabolic adaptation relevant to human disease.
Keywords: BioID; IGF2BP2; RNA-binding proteins; Z-disc; actinin; mRNA localization; oxidative phosphorylation; protein-protein interactions; quantitative proteomics; sarcomere.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Aye TT, Soni S, van Veen TA, van der Heyden MA, Cappadona S, Varro A, de Weger RA, de Jonge N, Vos MA, Heck AJ, and Scholten A (2012). Reorganized PKA-AKAP associations in the failing human heart. J. Mol. Cell. Cardiol 52, 511–518. - PubMed
-
- Benjamini Y, Krieger AM, and Yekutieli D (2006). Adaptive linear step-up procedures that control the false discovery rate. Biometrika 93, 491–507.
-
- Beqqali A, Monshouwer-Kloots J, Monteiro R, Welling M, Bakkers J, Ehler E, Verkleij A, Mummery C, and Passier R (2010). CHAP is a newly identified Z-disc protein essential for heart and skeletal muscle function. J. Cell Sci 123, 1141–1150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
