

Mutations in the transcription factor FOXO1 mimic positive selection signals to promote germinal center B cell expansion and lymphomagenesis
- PMID: 34380064
- PMCID: PMC8475267
- DOI: 10.1016/j.immuni.2021.07.009
Mutations in the transcription factor FOXO1 mimic positive selection signals to promote germinal center B cell expansion and lymphomagenesis
Abstract
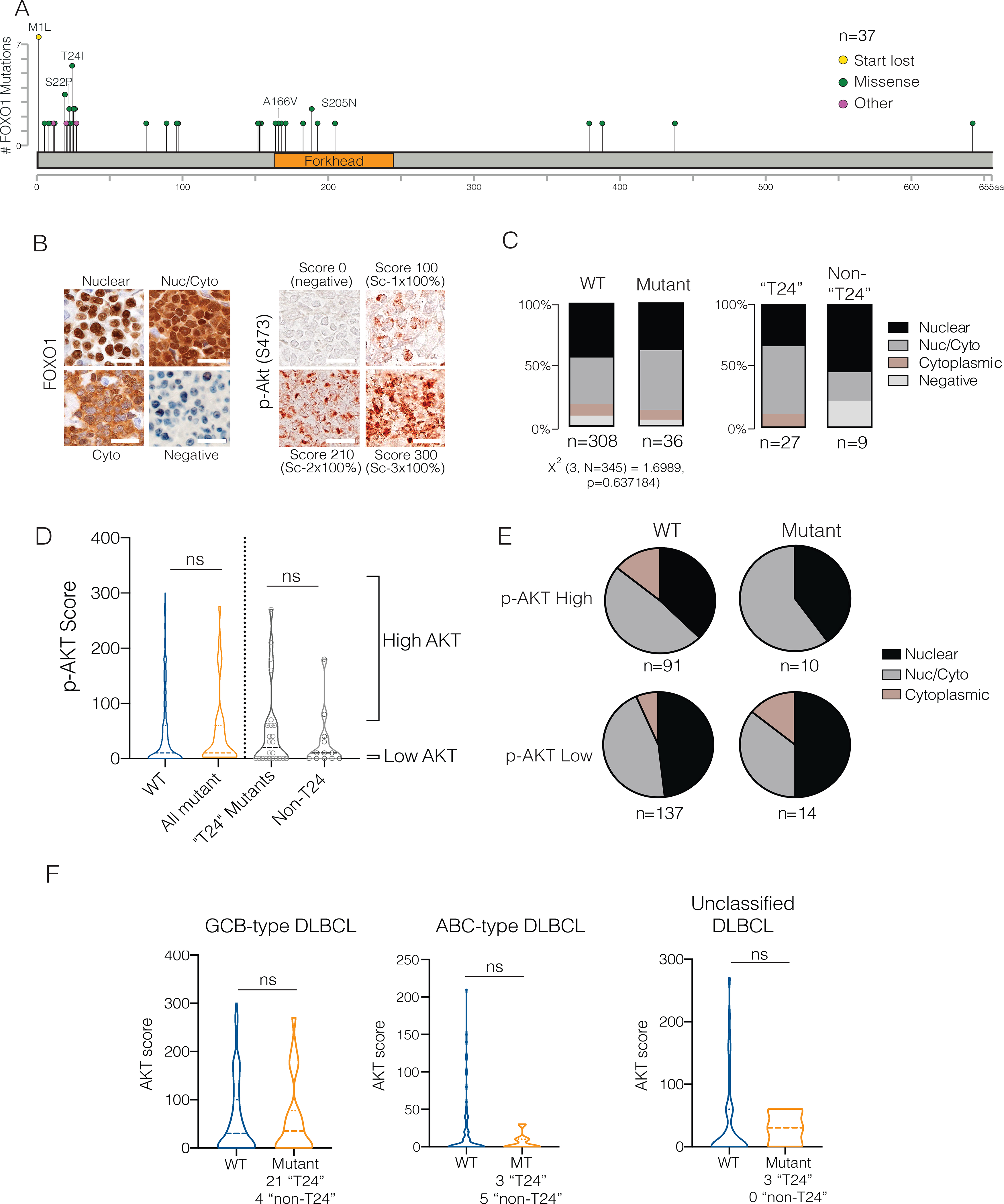
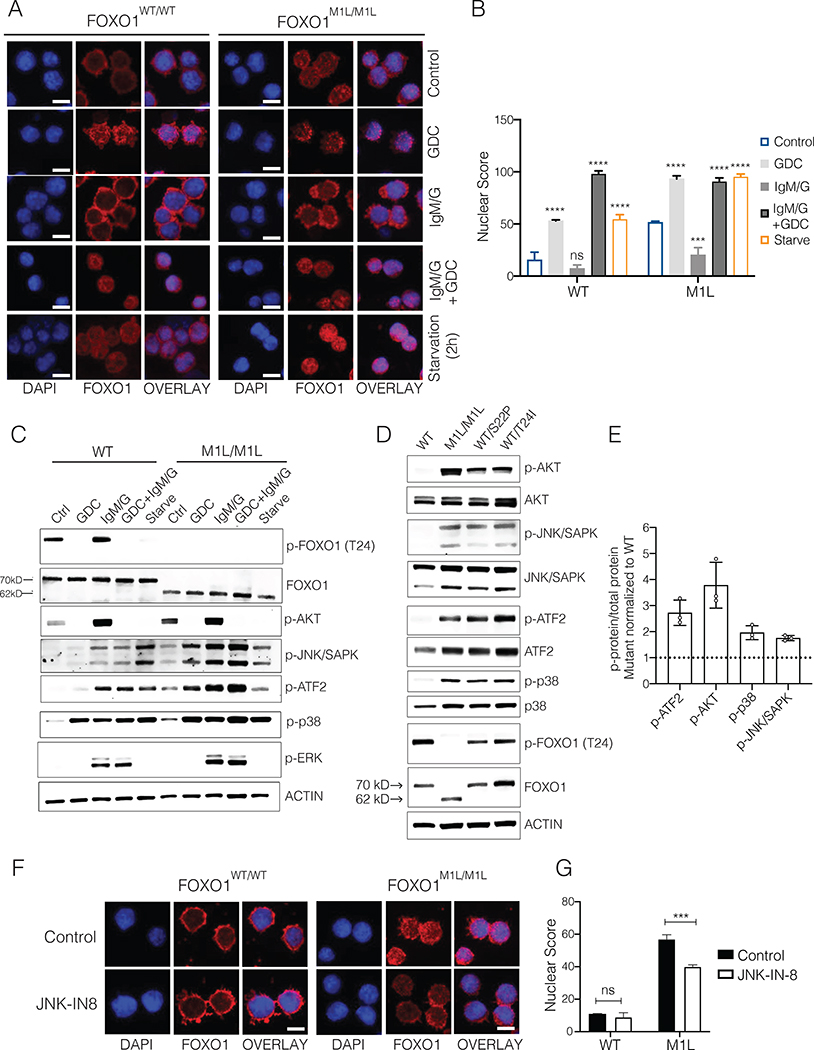
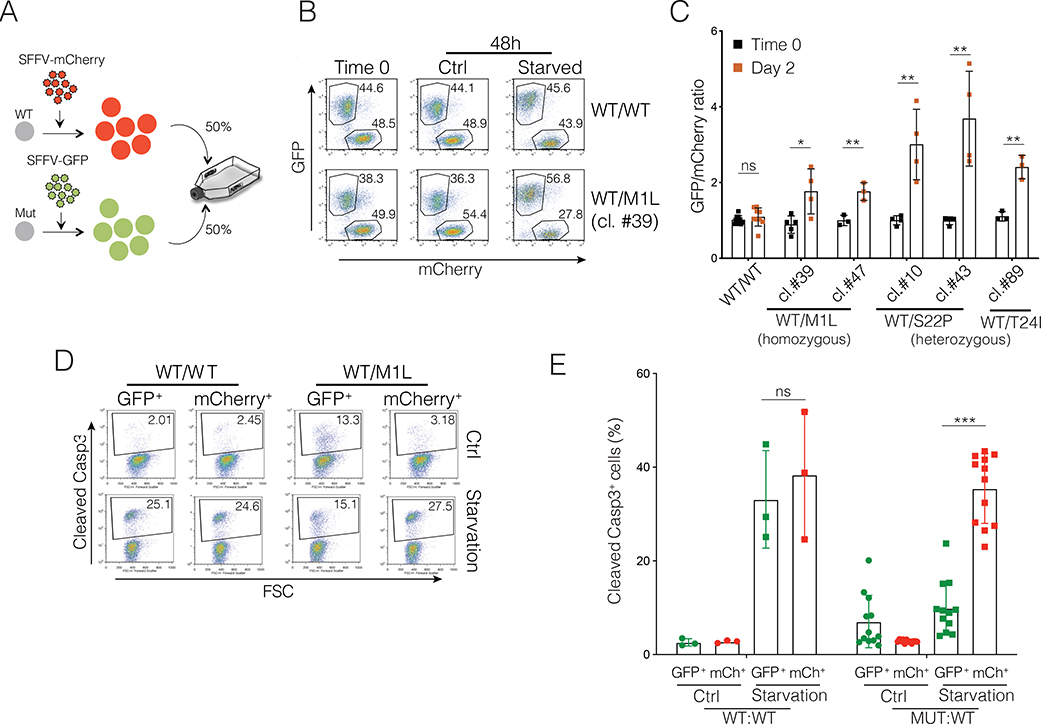
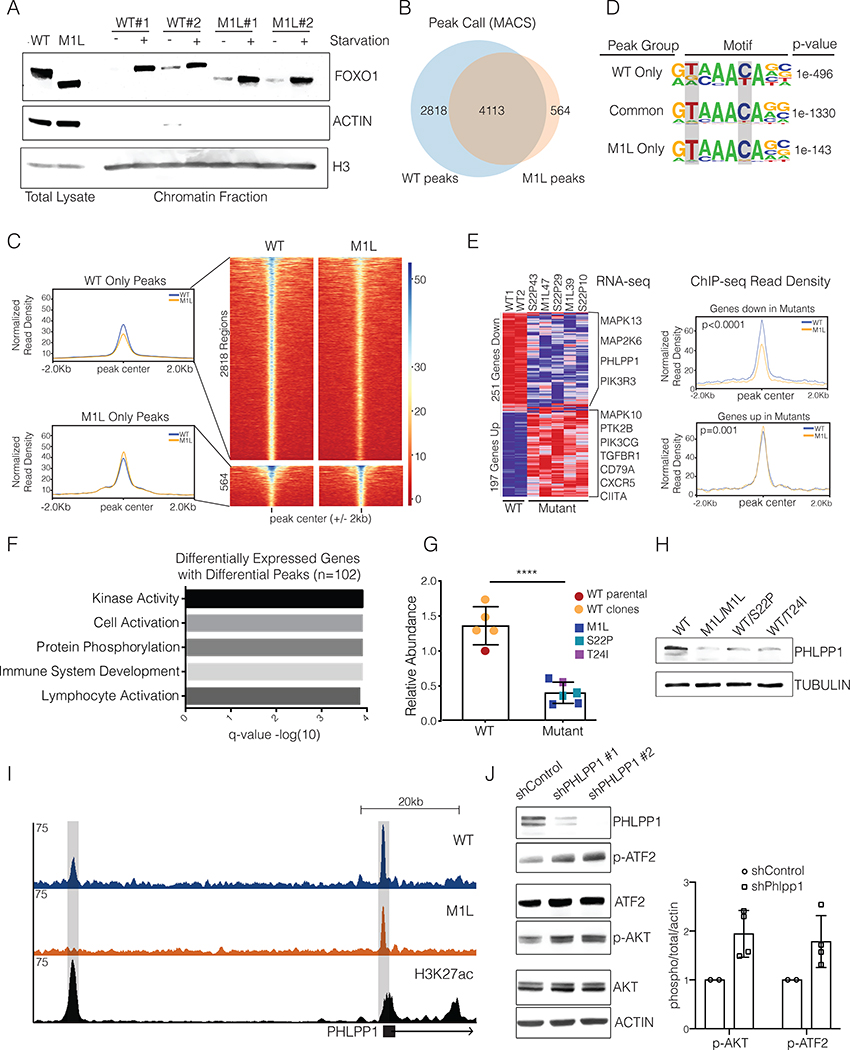
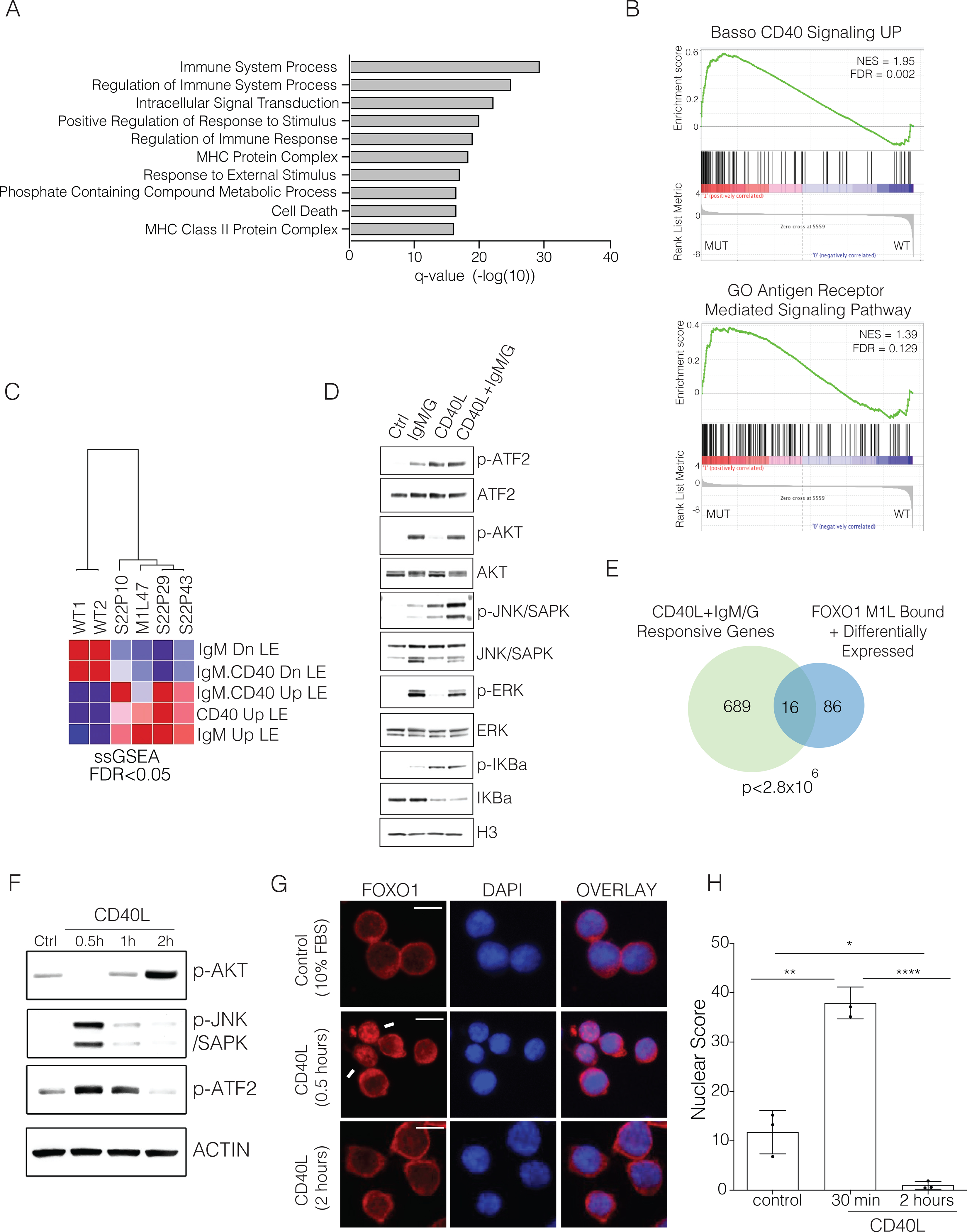
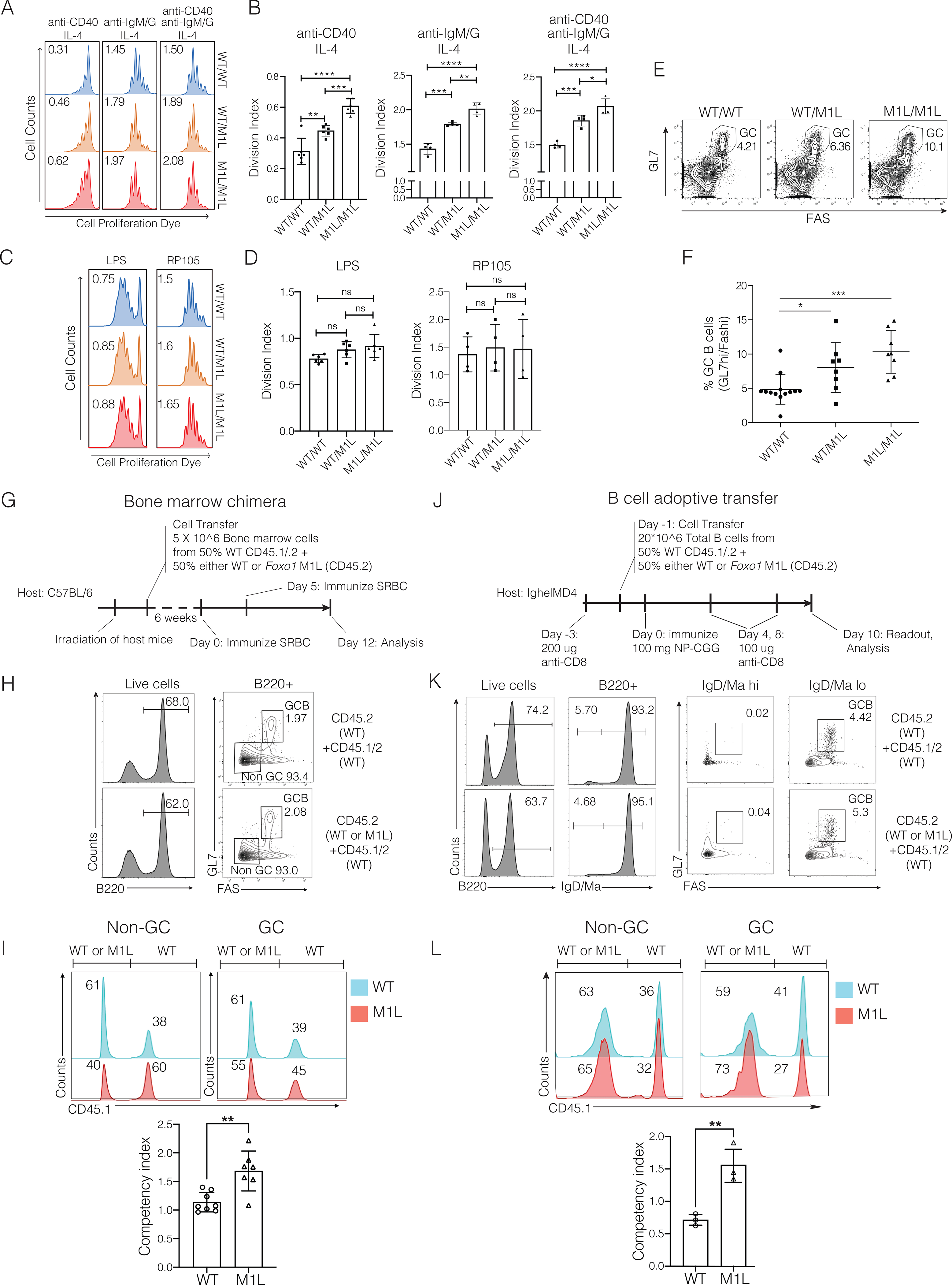
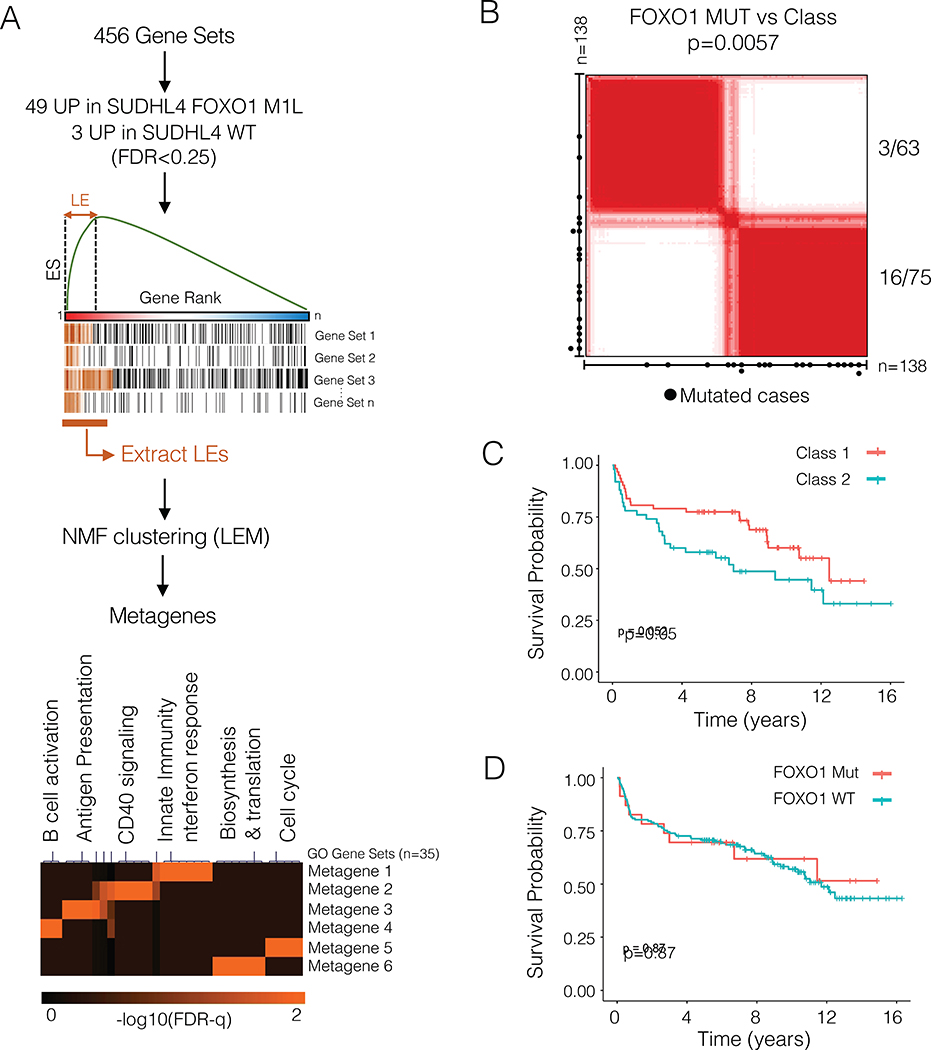
The transcription factor forkhead box O1 (FOXO1), which instructs the dark zone program to direct germinal center (GC) polarity, is typically inactivated by phosphatidylinositol 3-kinase (PI3K) signals. Here, we investigated how FOXO1 mutations targeting this regulatory axis in GC-derived B cell non-Hodgkin lymphomas (B-NHLs) contribute to lymphomagenesis. Examination of primary B-NHL tissues revealed that FOXO1 mutations and PI3K pathway activity were not directly correlated. Human B cell lines bearing FOXO1 mutations exhibited hyperactivation of PI3K and Stress-activated protein kinase (SAPK)/Jun amino-terminal kinase (JNK) signaling, and increased cell survival under stress conditions as a result of alterations in FOXO1 transcriptional affinities and activation of transcriptional programs characteristic of GC-positive selection. When modeled in mice, FOXO1 mutations conferred competitive advantage to B cells in response to key T-dependent immune signals, disrupting GC homeostasis. FOXO1 mutant transcriptional signatures were prevalent in human B-NHL and predicted poor clinical outcomes. Thus, rather than enforcing FOXO1 constitutive activity, FOXO1 mutations enable co-option of GC-positive selection programs during the pathogenesis of GC-derived lymphomas.
Keywords: B cell; CD40; FOXO1; JNK; PI3K; germinal center; mouse model; mutation; non-Hodgkin lymphoma; positive selection.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Nuclear FOXO1 promotes lymphomagenesis in germinal center B cells.Blood. 2018 Dec 20;132(25):2670-2683. doi: 10.1182/blood-2018-06-856203. Epub 2018 Oct 17. Blood. 2018. PMID: 30333121
-
The FOXO1 Transcription Factor Instructs the Germinal Center Dark Zone Program.Immunity. 2015 Dec 15;43(6):1064-74. doi: 10.1016/j.immuni.2015.10.015. Epub 2015 Nov 24. Immunity. 2015. PMID: 26620759
-
PI3 Kinase and FOXO1 Transcription Factor Activity Differentially Control B Cells in the Germinal Center Light and Dark Zones.Immunity. 2015 Dec 15;43(6):1075-86. doi: 10.1016/j.immuni.2015.10.021. Epub 2015 Nov 24. Immunity. 2015. PMID: 26620760
-
FOXO1 transcription factor: a critical effector of the PI3K-AKT axis in B-cell development.Int Rev Immunol. 2014 Mar;33(2):146-57. doi: 10.3109/08830185.2014.885022. Epub 2014 Feb 20. Int Rev Immunol. 2014. PMID: 24552152 Review.
-
Human B Lymphomas Reveal Their Secrets Through Genetic Mouse Models.Front Immunol. 2021 Jul 16;12:683597. doi: 10.3389/fimmu.2021.683597. eCollection 2021. Front Immunol. 2021. PMID: 34335584 Free PMC article. Review.
Cited by
-
STAT3 signaling in B cells controls germinal center zone organization and recycling.Cell Rep. 2023 May 30;42(5):112512. doi: 10.1016/j.celrep.2023.112512. Epub 2023 May 16. Cell Rep. 2023. PMID: 37200190 Free PMC article.
-
The discrete roles of individual FOXO transcription factor family members in B-cell malignancies.Front Immunol. 2023 May 18;14:1179101. doi: 10.3389/fimmu.2023.1179101. eCollection 2023. Front Immunol. 2023. PMID: 37275916 Free PMC article. Review.
-
The transcriptional program during germinal center reaction - a close view at GC B cells, Tfh cells and Tfr cells.Front Immunol. 2023 Feb 3;14:1125503. doi: 10.3389/fimmu.2023.1125503. eCollection 2023. Front Immunol. 2023. PMID: 36817488 Free PMC article. Review.
-
Stratified Medicine Pediatrics: Cell-Free DNA and Serial Tumor Sequencing Identifies Subtype-Specific Cancer Evolution and Epigenetic States.Cancer Discov. 2025 Apr 2;15(4):717-732. doi: 10.1158/2159-8290.CD-24-0916. Cancer Discov. 2025. PMID: 39693475 Free PMC article.
-
Mutant FOXO1 controls an oncogenic network via enhancer accessibility.Cell Genom. 2024 Apr 10;4(4):100537. doi: 10.1016/j.xgen.2024.100537. Cell Genom. 2024. PMID: 38604128 Free PMC article.
References
-
- Aggarwal M, Sanchez-Beato M, Gomez-Lopez G, Al-Shahrour F, Martínez N, Rodríguez A, Ruiz-Ballesteros E, Camacho FI, Perez-Rosado A, de la Cueva P, et al. (2009). Functional signatures identified in B-cell non-Hodgkin lymphoma profiles. Leuk Lymphoma 50, 1699–1708. - PubMed
-
- Alizadeh AA, Eisen MB, Davis RE, Ma C, Lossos IS, Rosenwald A, Boldrick JC, Sabet H, Tran T, Yu X, et al. (2000). Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 403, 503–511. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
