

A broadly active fucosyltransferase LmjFUT1 whose mitochondrial localization and activity are essential in parasitic Leishmania
- PMID: 34385330
- PMCID: PMC8379939
- DOI: 10.1073/pnas.2108963118
A broadly active fucosyltransferase LmjFUT1 whose mitochondrial localization and activity are essential in parasitic Leishmania
Abstract
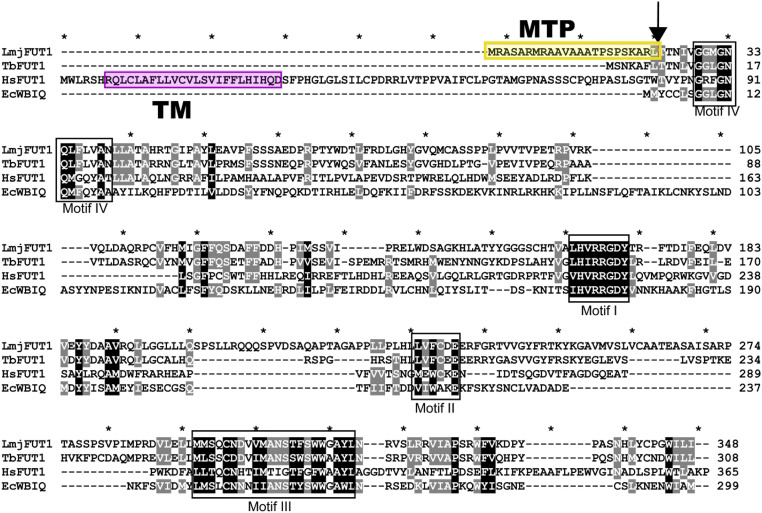
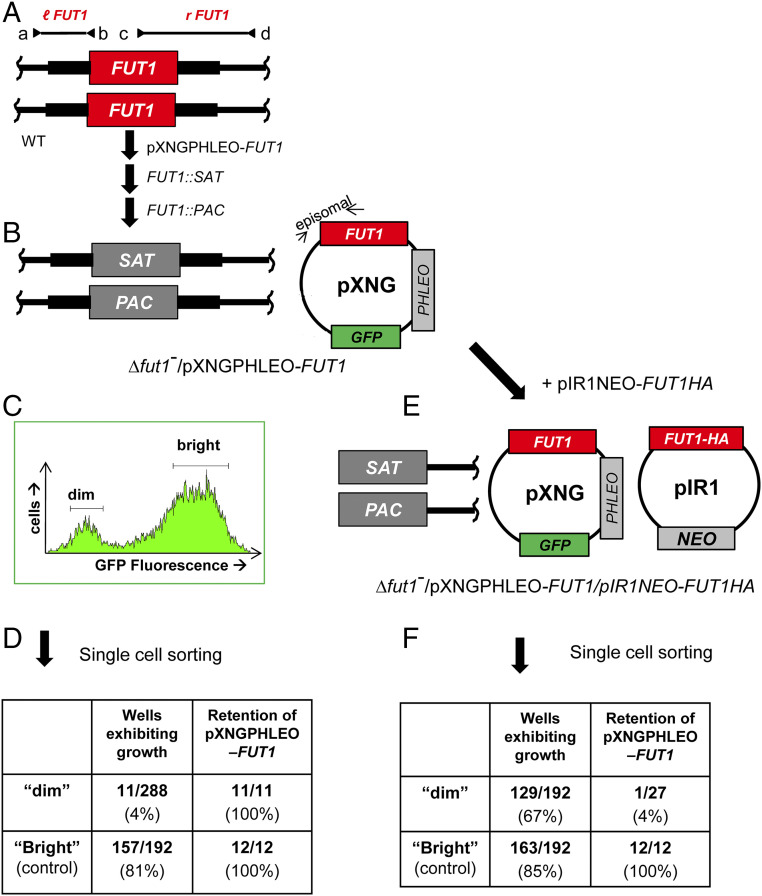
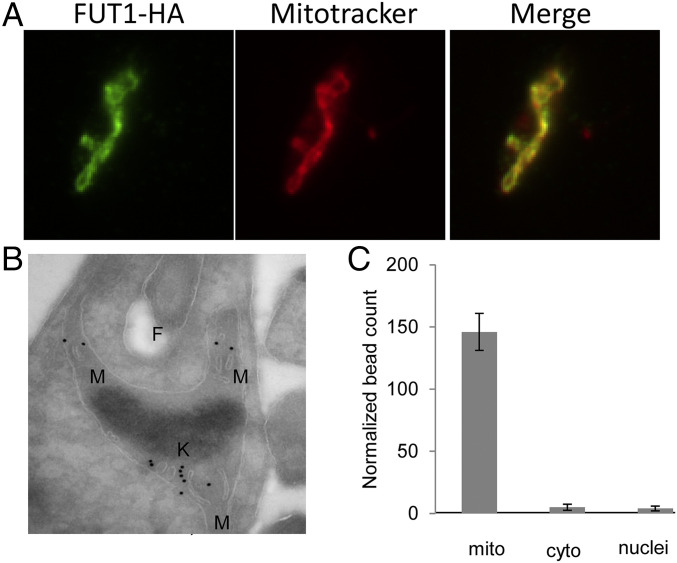
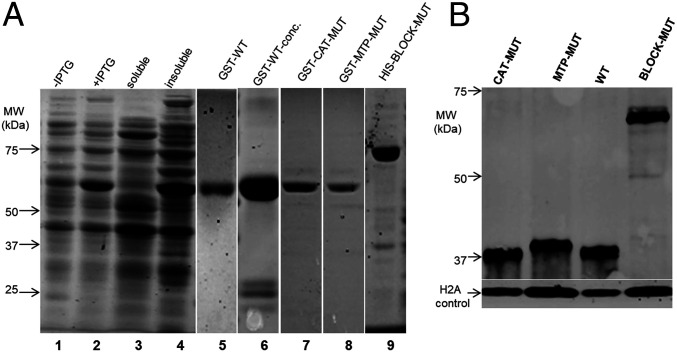
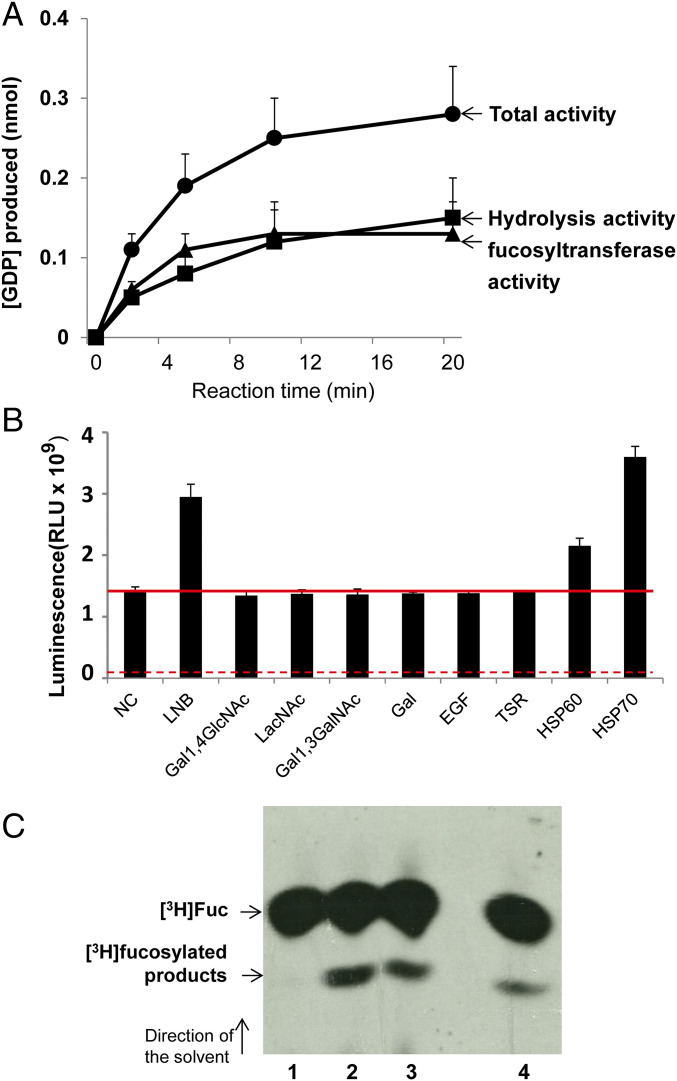
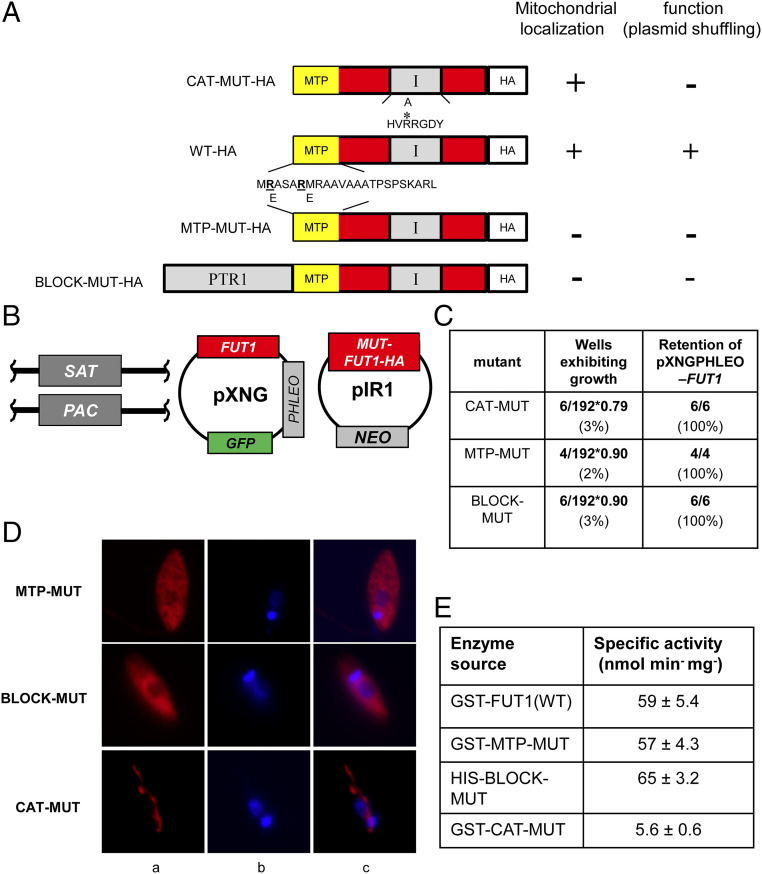
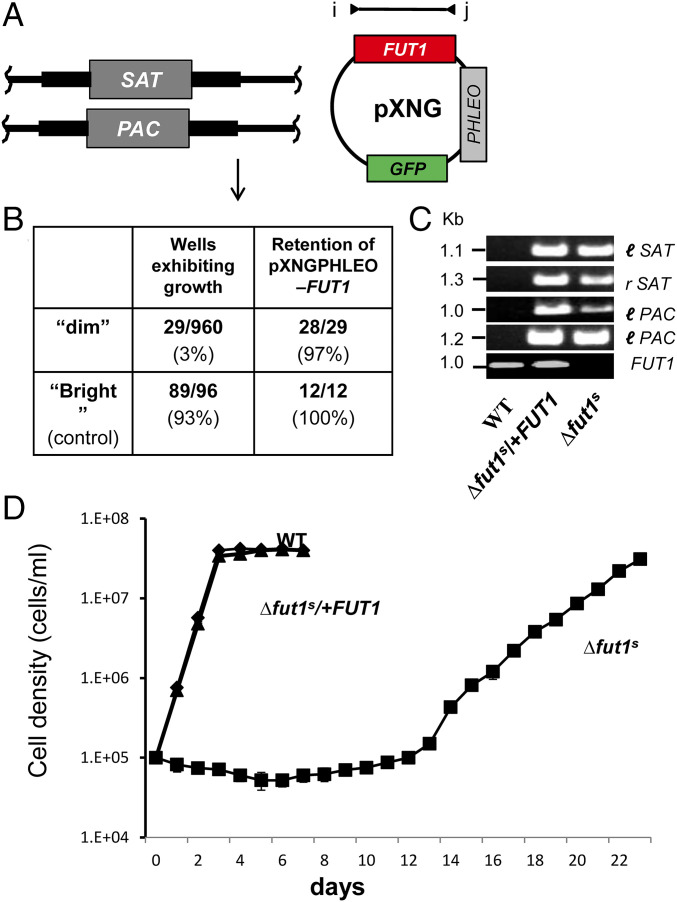
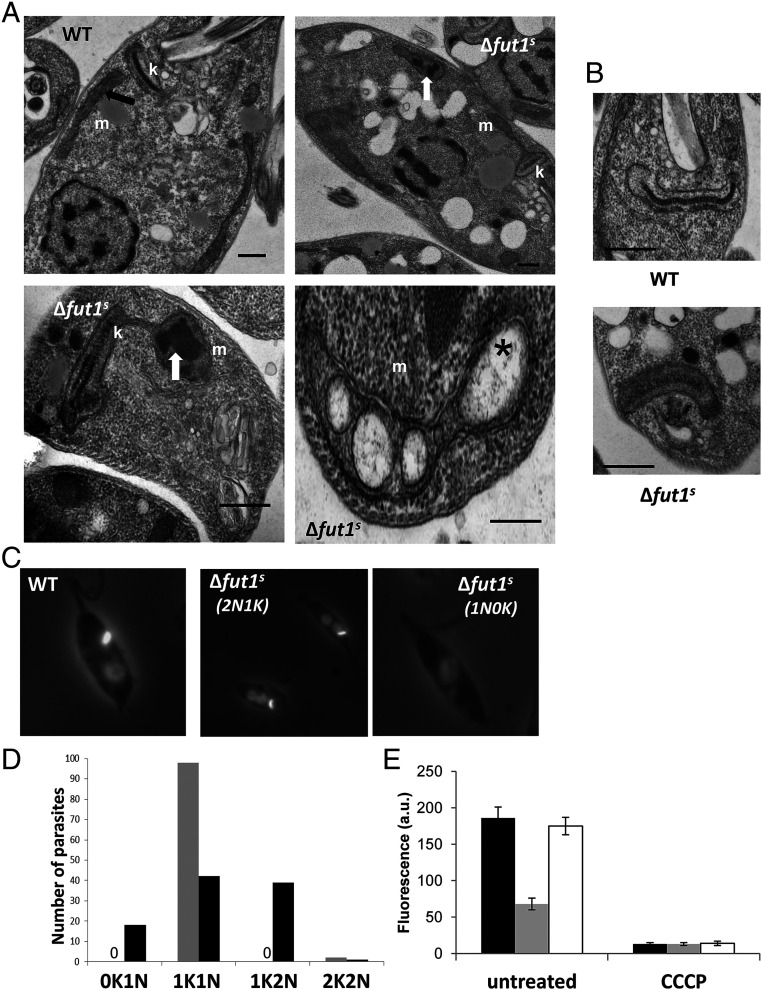
Glycoconjugates play major roles in the infectious cycle of the trypanosomatid parasite Leishmania While GDP-Fucose synthesis is essential, fucosylated glycoconjugates have not been reported in Leishmania major [H. Guo et al., J. Biol. Chem. 292, 10696-10708 (2017)]. Four predicted fucosyltransferases appear conventionally targeted to the secretory pathway; SCA1/2 play a role in side-chain modifications of lipophosphoglycan, while gene deletion studies here showed that FUT2 and SCAL were not essential. Unlike most eukaryotic glycosyltransferases, the predicted α 1-2 fucosyltransferase encoded by FUT1 localized to the mitochondrion. A quantitative "plasmid segregation" assay, expressing FUT1 from the multicopy episomal pXNG vector in a chromosomal null ∆fut1- background, established that FUT1 is essential. Similarly, "plasmid shuffling" confirmed that both enzymatic activity and mitochondrial localization were required for viability, comparing import-blocked or catalytically inactive enzymes, respectively. Enzymatic assays of tagged proteins expressed in vivo or of purified recombinant FUT1 showed it had a broad fucosyltransferase activity including glycan and peptide substrates. Unexpectedly, a single rare ∆fut1- segregant (∆fut1s ) was obtained in rich media, which showed severe growth defects accompanied by mitochondrial dysfunction and loss, all of which were restored upon FUT1 reexpression. Thus, FUT1 along with the similar Trypanosoma brucei enzyme TbFUT1 [G. Bandini et al., bioRxiv, https://www.biorxiv.org/content/10.1101/726117v2 (2021)] joins the eukaryotic O-GlcNAc transferase isoform as one of the few glycosyltransferases acting within the mitochondrion. Trypanosomatid mitochondrial FUT1s may offer a facile system for probing mitochondrial glycosylation in a simple setting, and their essentiality for normal growth and mitochondrial function renders it an attractive target for chemotherapy of these serious human pathogens.
Keywords: chemotherapy; fucose; glycobiology; glycosyltransferase; trypanosomatid protozoan parasites.
Conflict of interest statement
Competing interest statement: One of the reviewers (G.W.H.) and one of the authors (M.A.J.F.) are coauthors on chapter 12 in Essentials of Glycobiology (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, ed. 3, 2015–2017).
Figures
References
-
- Bañuls A. L., et al. ., Clinical pleiomorphism in human leishmaniases, with special mention of asymptomatic infection. Clin. Microbiol. Infect. 17, 1451–1461 (2011). - PubMed
-
- Naderer T., McConville M. J., Intracellular growth and pathogenesis of Leishmania parasites. Essays Biochem. 51, 81–95 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
