

Morphometric analysis of astrocytes in vocal production circuits of common marmoset (Callithrix jacchus)
- PMID: 34387357
- PMCID: PMC8716418
- DOI: 10.1002/cne.25230
Morphometric analysis of astrocytes in vocal production circuits of common marmoset (Callithrix jacchus)
Abstract
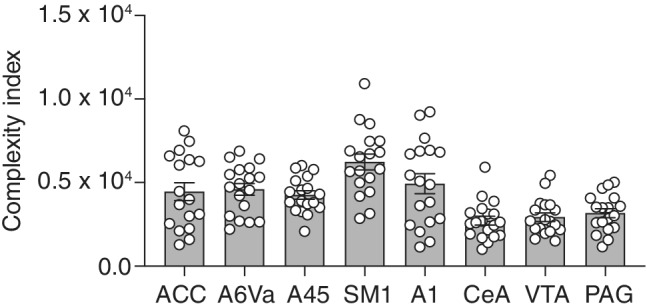
Astrocytes, the star-shaped glial cells, are the most abundant non-neuronal cell population in the central nervous system. They play a key role in modulating activities of neural networks, including those involved in complex motor behaviors. Common marmosets (Callithrix jacchus), the most vocal non-human primate (NHP), have been used to study the physiology of vocalization and social vocal production. However, the neural circuitry involved in vocal production is not fully understood. In addition, even less is known about the involvement of astrocytes in this circuit. To understand the role, that astrocytes may play in the complex behavior of vocalization, the initial step may be to study their structural properties in the cortical and subcortical regions that are known to be involved in vocalization. Here, in the common marmoset, we identify all astrocytic subtypes seen in other primate's brains, including intralaminar astrocytes. In addition, we reveal detailed structural characteristics of astrocytes and perform morphometric analysis of astrocytes residing in the cortex and midbrain regions that are associated with vocal production. We found that cortical astrocytes in these regions illustrate a higher level of complexity when compared to those in the midbrain. We hypothesize that this complexity that is expressed in cortical astrocytes may reflect their functions to meet the metabolic/structural needs of these regions.
Keywords: GFAP; astrocytes; cerebral cortex; common marmoset; glia; midbrain; vocalization.
Published 2021. This article is a U.S. Government work and is in the public domain in the USA.
Conflict of interest statement
The authors declare no competing financial interests.
Figures








References
-
- Althammer, F. , Ferreira‐Neto, H. C. , Rubaharan, M. , Roy, R. K. , Patel, A. A. , Murphy, A. , Cox, D. N. , & Stern, J. E. (2020). Three‐dimensional morphometric analysis reveals time‐dependent structural changes in microglia and astrocytes in the central amygdala and hypothalamic paraventricular nucleus of heart failure rats. Journal of Neuroinflammation, 17, 221. - PMC - PubMed
-
- Anderson, C. M. , & Swanson, R. A. (2000). Astrocyte glutamate transport: Review of properties, regulation, and physiological functions. Glia, 32, 1–14. - PubMed
-
- Angelova, P. R. , Kasymov, V. , Christie, I. , Sheikhbahaei, S. , Turovsky, E. , Marina, N. , Korsak, A. , Zwicker, J. , Teschemacher, A. G. , Ackland, G. L. , Funk, G. D. , Kasparov, S. , Abramov, A. Y. , & Gourine, A. V. (2015). Functional oxygen sensitivity of astrocytes. The Journal of Neuroscience, 35, 10460–10473. - PMC - PubMed
-
- Bakola, S. , Burman, K. J. , & Rosa, M. G. P. (2015). The cortical motor system of the marmoset monkey (Callithrix jacchus). Neuroscience Research, 93, 72–81. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
