

Developmental and temporal characteristics of clonal sperm mosaicism
- PMID: 34388390
- PMCID: PMC8496133
- DOI: 10.1016/j.cell.2021.07.024
Developmental and temporal characteristics of clonal sperm mosaicism
Abstract
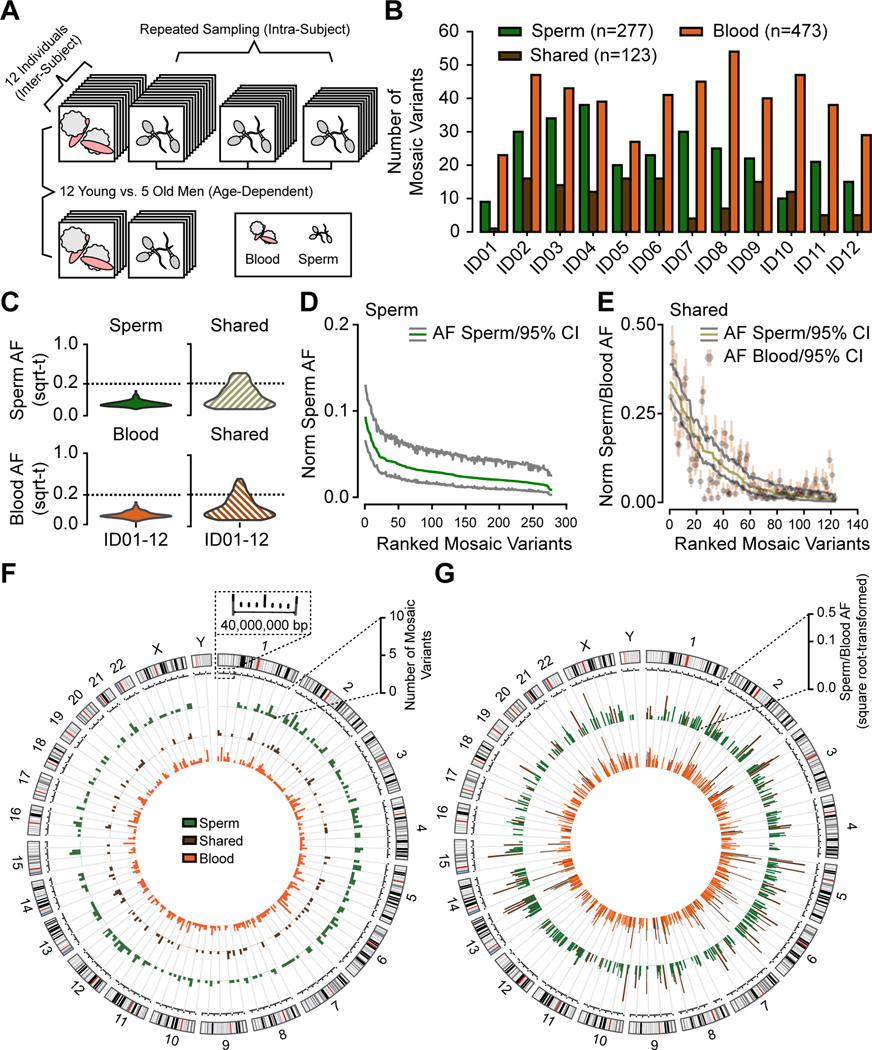
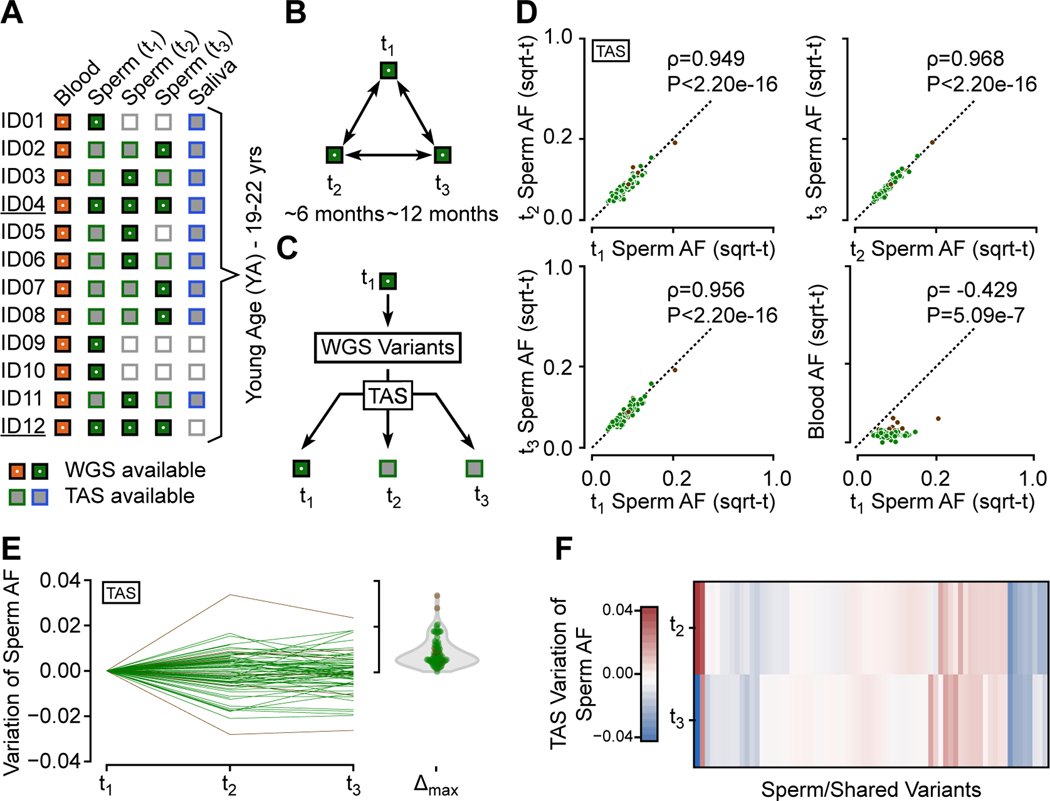
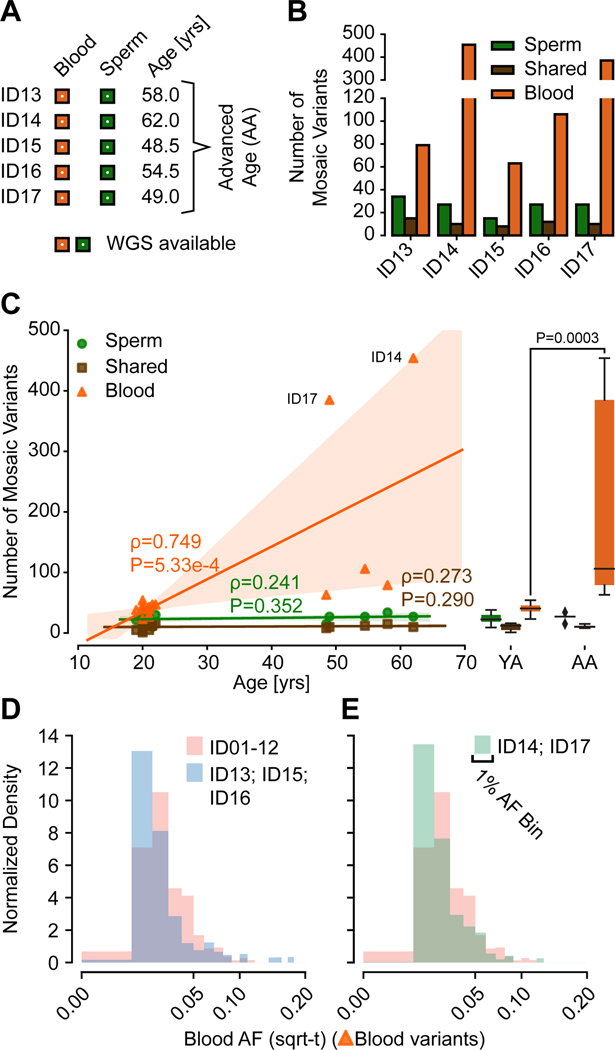
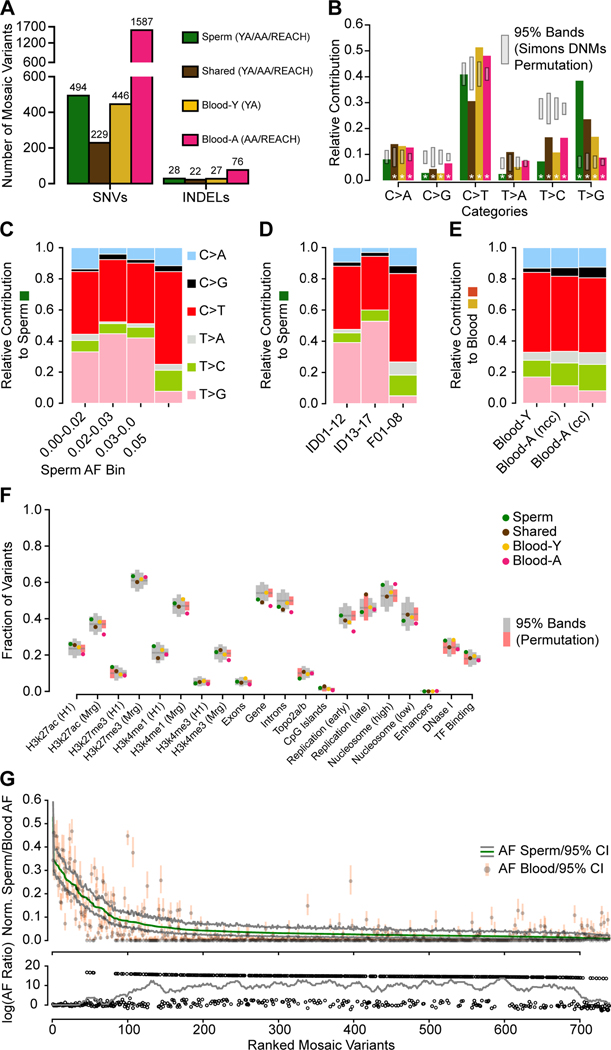
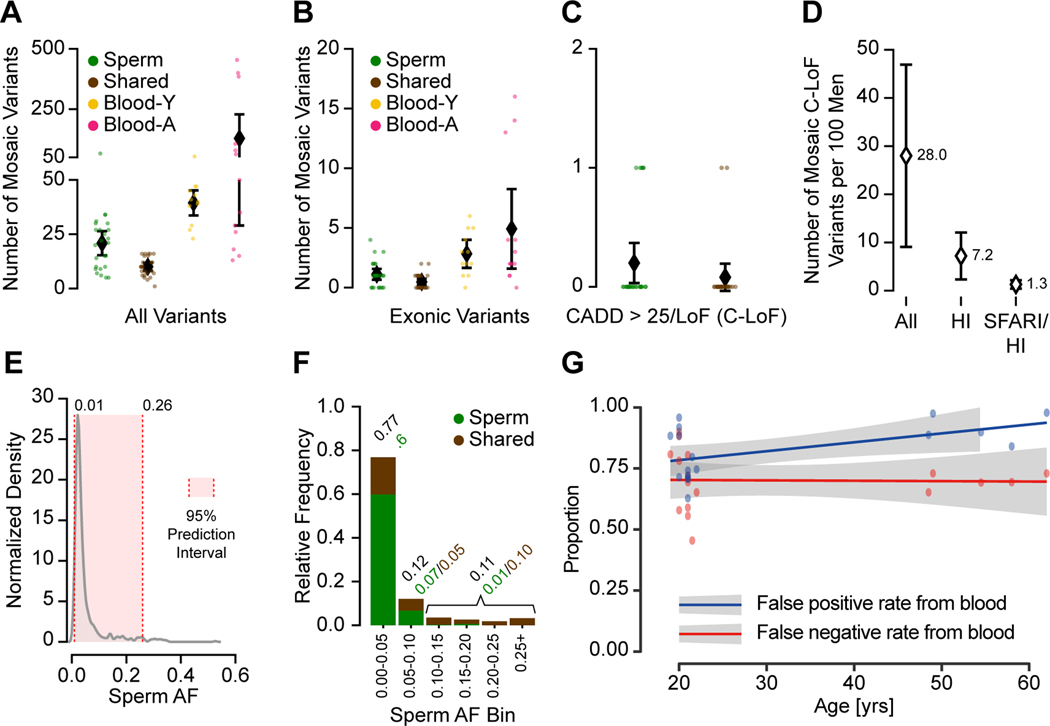
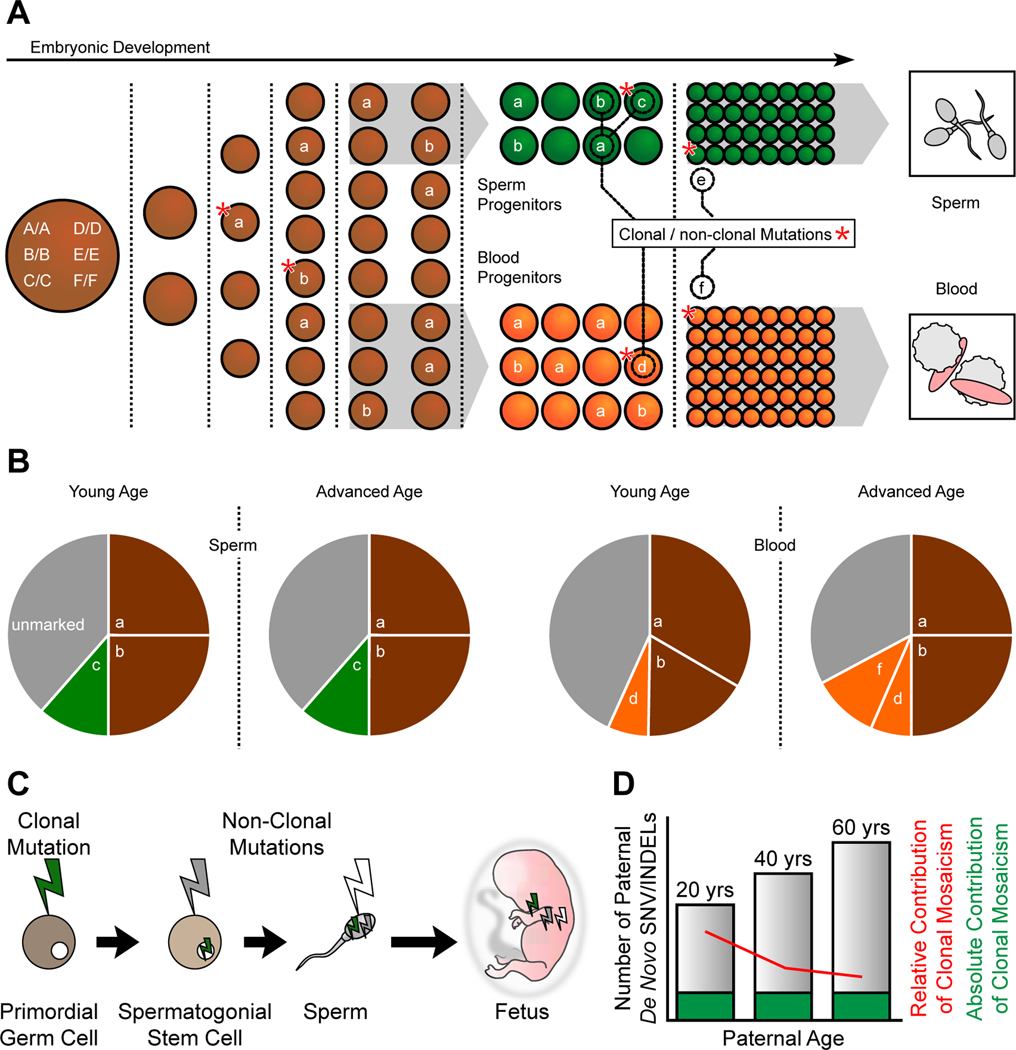
Throughout development and aging, human cells accumulate mutations resulting in genomic mosaicism and genetic diversity at the cellular level. Mosaic mutations present in the gonads can affect both the individual and the offspring and subsequent generations. Here, we explore patterns and temporal stability of clonal mosaic mutations in male gonads by sequencing ejaculated sperm. Through 300× whole-genome sequencing of blood and sperm from healthy men, we find each ejaculate carries on average 33.3 ± 12.1 (mean ± SD) clonal mosaic variants, nearly all of which are detected in serial sampling, with the majority absent from sampled somal tissues. Their temporal stability and mutational signature suggest origins during embryonic development from a largely immutable stem cell niche. Clonal mosaicism likely contributes a transmissible, predicted pathogenic exonic variant for 1 in 15 men, representing a life-long threat of transmission for these individuals and a significant burden on human population health.
Keywords: autism spectrum disorder; clonal mosaicism; congenital disorders; de novo mutation; embryogenesis; mutational signature; somatic; sperm; transmission risk.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.W.B., D.A., K.N.J., J.S., and J.G.G. are inventors on a patent (PCT/US2018/024878, WO2018183525A1) filed by University of California, San Diego that is titled “Methods for assessing risk of or diagnosing genetic defects by identifying de novo mutations or somatic mosaic variants in sperm or somatic tissues”.
Figures
References
-
- Alankarage D, Ip E, Szot JO, Munro J, Blue GM, Harrison K, Cuny H, Enriquez A, Troup M, Humphreys DT, et al. (2019). Identification of clinically actionable variants from genome sequencing of families with congenital heart disease. Genet Med 21, 1111–1120. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
