

A systems-level analysis highlights microglial activation as a modifying factor in common epilepsies
- PMID: 34388852
- PMCID: PMC8983060
- DOI: 10.1111/nan.12758
A systems-level analysis highlights microglial activation as a modifying factor in common epilepsies
Abstract
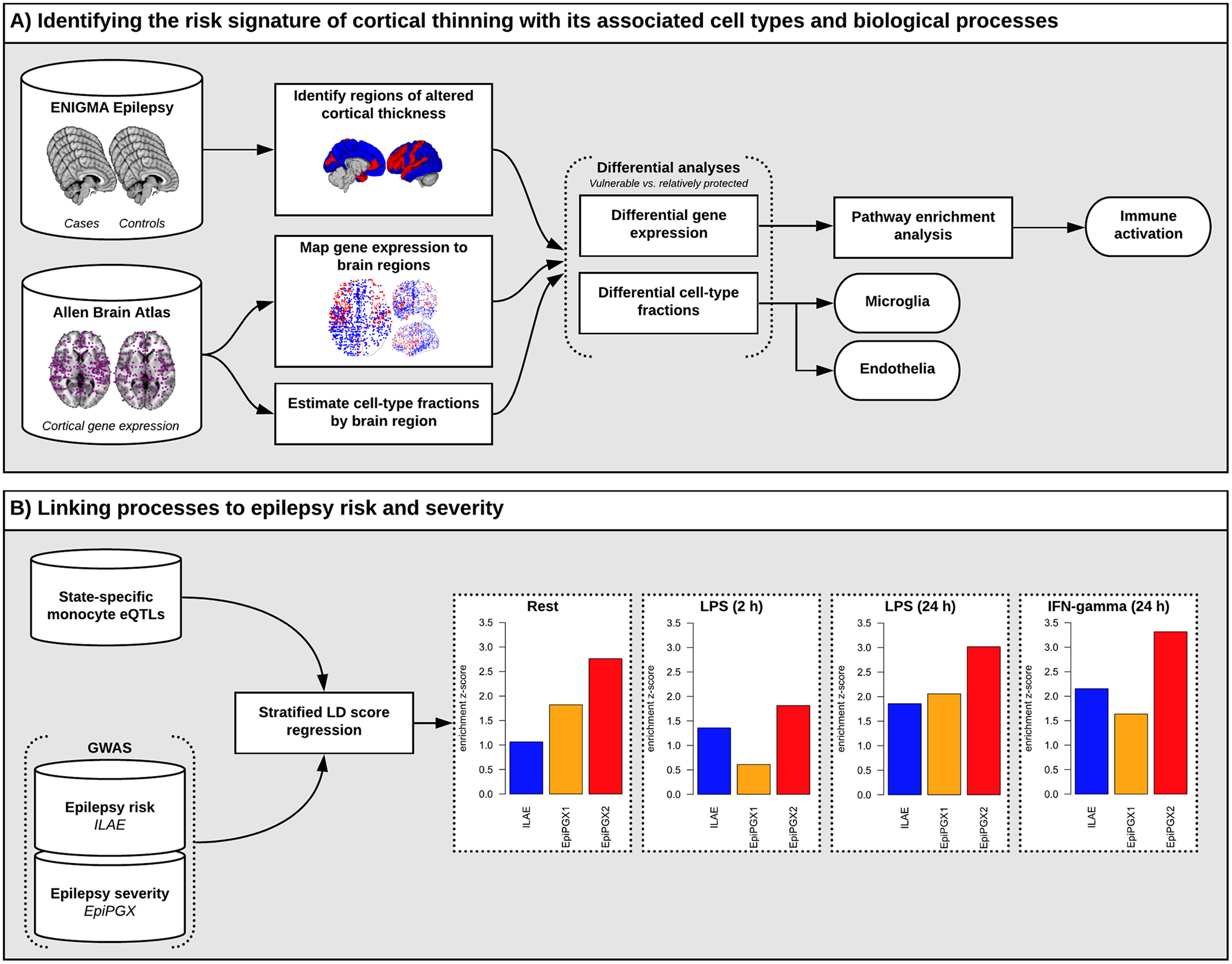
Aims: The causes of distinct patterns of reduced cortical thickness in the common human epilepsies, detectable on neuroimaging and with important clinical consequences, are unknown. We investigated the underlying mechanisms of cortical thinning using a systems-level analysis.
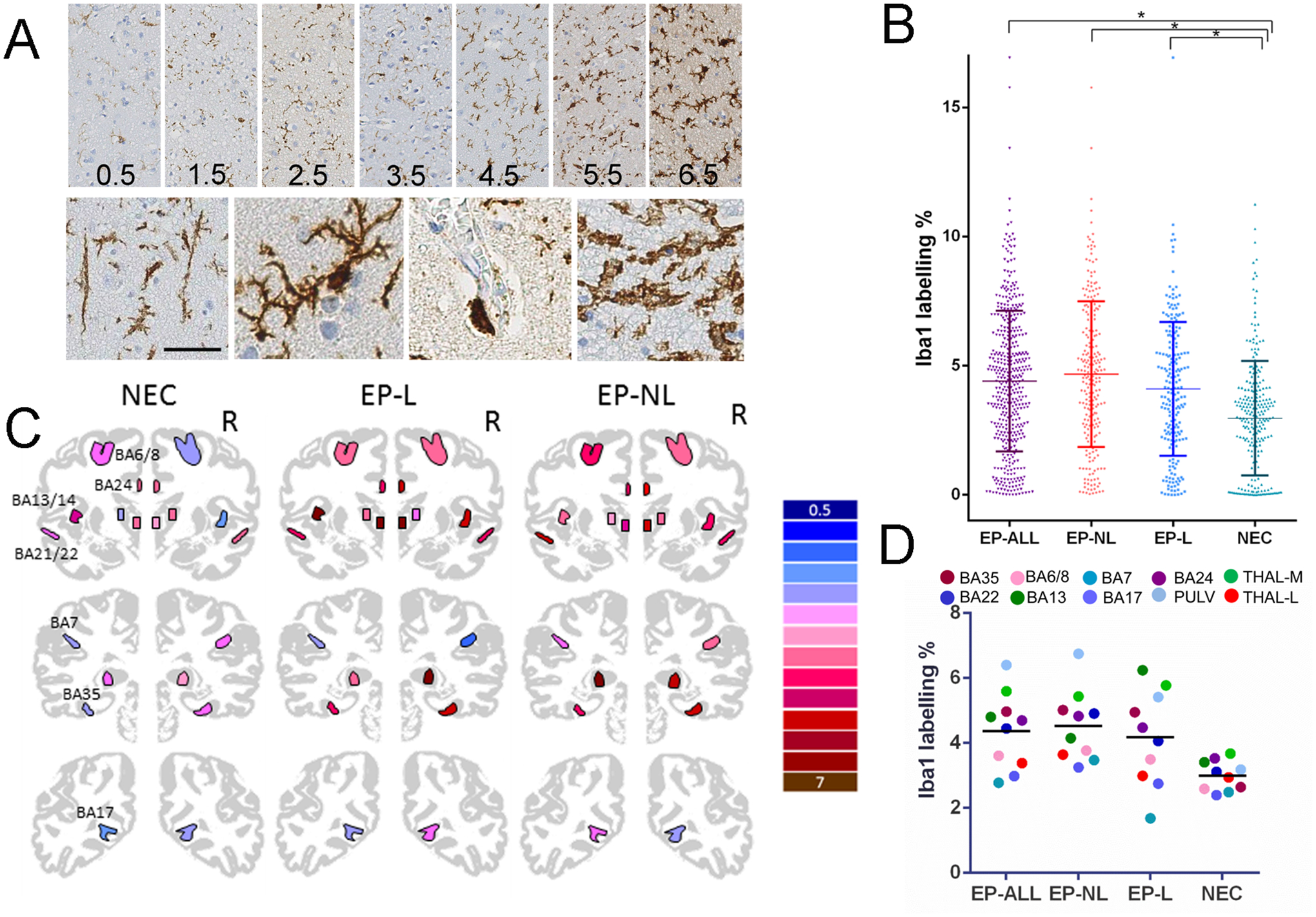
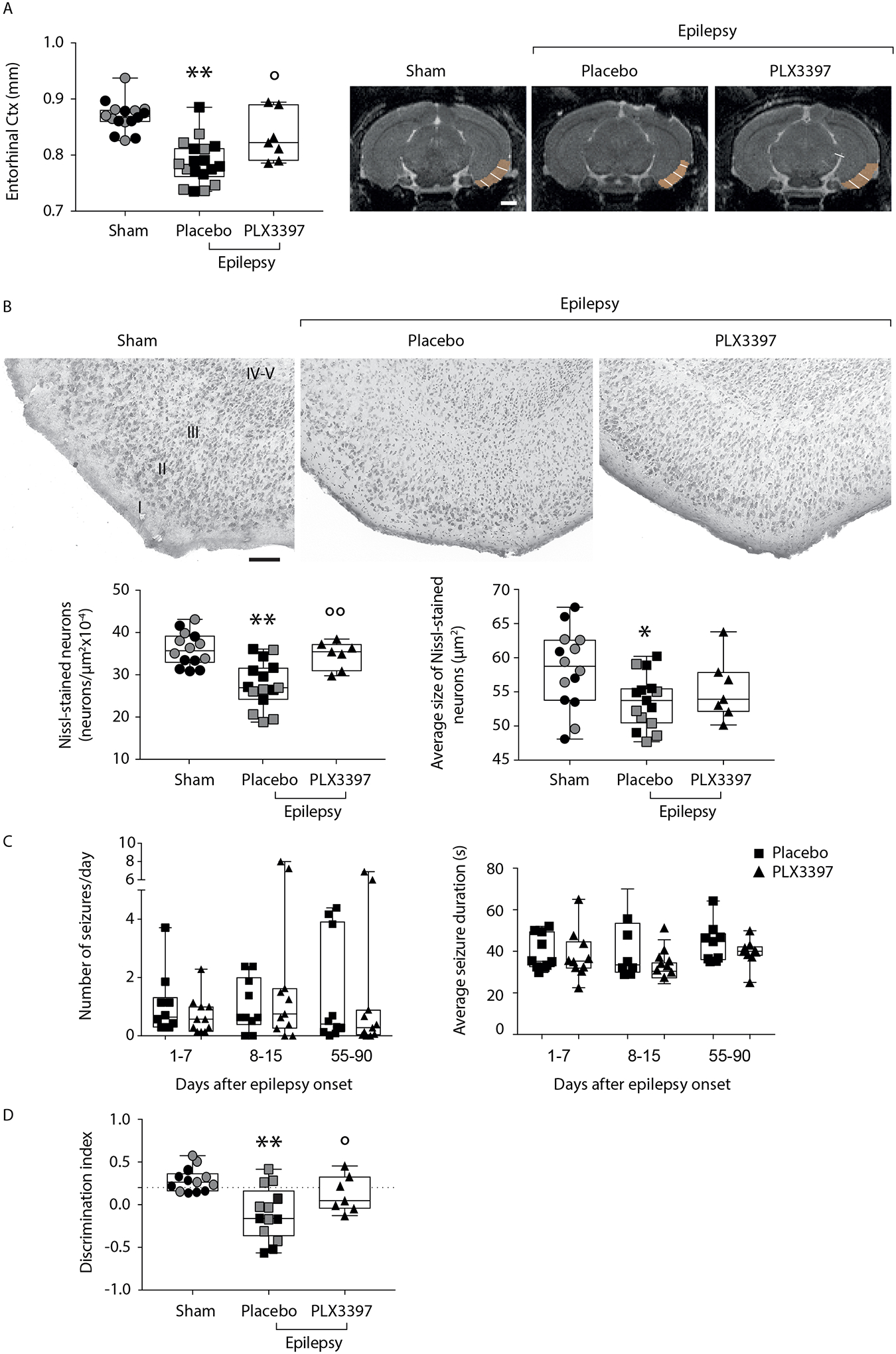
Methods: Imaging-based cortical structural maps from a large-scale epilepsy neuroimaging study were overlaid with highly spatially resolved human brain gene expression data from the Allen Human Brain Atlas. Cell-type deconvolution, differential expression analysis and cell-type enrichment analyses were used to identify differences in cell-type distribution. These differences were followed up in post-mortem brain tissue from humans with epilepsy using Iba1 immunolabelling. Furthermore, to investigate a causal effect in cortical thinning, cell-type-specific depletion was used in a murine model of acquired epilepsy.
Results: We identified elevated fractions of microglia and endothelial cells in regions of reduced cortical thickness. Differentially expressed genes showed enrichment for microglial markers and, in particular, activated microglial states. Analysis of post-mortem brain tissue from humans with epilepsy confirmed excess activated microglia. In the murine model, transient depletion of activated microglia during the early phase of the disease development prevented cortical thinning and neuronal cell loss in the temporal cortex. Although the development of chronic seizures was unaffected, the epileptic mice with early depletion of activated microglia did not develop deficits in a non-spatial memory test seen in epileptic mice not depleted of microglia.
Conclusions: These convergent data strongly implicate activated microglia in cortical thinning, representing a new dimension for concern and disease modification in the epilepsies, potentially distinct from seizure control.
Keywords: MRI; cortical thinning; gene expression; post mortem.
© 2021 The Authors. Neuropathology and Applied Neurobiology published by John Wiley & Sons Ltd on behalf of British Neuropathological Society.
Conflict of interest statement
Conflict of Interest statements
C.D.W. is now an employee of Biogen Research and Early Development (Cambridge, Massachusetts, 02142, USA).
Figures
Comment in
-
Cortical thinning in epilepsy is linked to microglial activation.Nat Rev Neurol. 2021 Oct;17(10):595. doi: 10.1038/s41582-021-00559-9. Nat Rev Neurol. 2021. PMID: 34475566 No abstract available.
References
-
- Orsini A, Zara F, Striano P. Recent advances in epilepsy genetics. Neurosci Lett. 2018;667:4–9. - PubMed
-
- Bell GS, Neligan A, Giavasi C, Keezer MR, Novy J, Peacock JL, et al. Outcome of seizures in the general population after 25 years: A prospective follow-up, observational cohort study. J Neurol Neurosurg Psychiatry. 2016;87(8):843–50. - PubMed
Publication types
MeSH terms
Grants and funding
- MR/N008324/1/Medical Research Council Clinician Scientist Fellowship
- R21 NS107739/NS/NINDS NIH HHS/United States
- MR/L016311/1/MRC_/Medical Research Council/United Kingdom
- U54 EB020403/EB/NIBIB NIH HHS/United States
- R01 NS065838/NH/NIH HHS/United States
- S10 OD023696/OD/NIH HHS/United States
- MR/N008324/1/MRC_/Medical Research Council/United Kingdom
- R01 EB015611/EB/NIBIB NIH HHS/United States
- R01 NS097719/NH/NIH HHS/United States
- R01 NS122827/NS/NINDS NIH HHS/United States
- R01 NS065838/NS/NINDS NIH HHS/United States
- R01 NS097719/NS/NINDS NIH HHS/United States
- U54 EB020403/NH/NIH HHS/United States
